

How do organisms adapt to climate change?
Chromosomal inversions in Drosophila subobscura: the case of Serbian populations

It is well known that chromosomal inversions in natural populations of the species Drosophila subobscura are adaptations to changes in the environment. It has been observed that inversions change in frequency over time and that they are so far following the pattern expected given the global warming of our planet. These results have been brought to light by our research on Serbian populations of D. subobscura. We found that inversion-related changes in chromosomal polymorphisms depended on the season of the year and in the long term (10-15 year periods) they followed what we would expect given global warming. Therefore, chromosomal inversions in this species are good indicators of climate change and can help us to study how organisms adapt to it.
Keywords: Drosophila subobscura, chromosome inversion, adaptation, natural selection, climate change.
Genetics and climate change
Living beings have the ability to adapt to environmental change. Adaptations are nothing more than characteristics of an organism that allow it to survive and reproduce. In order for this adaptive capacity to be useful, it must be able to move down generations, i.e. it needs to be hereditary. This is why genetics is so important when explaining how species face the challenge of an environmental deviation with such a reach and intensity as climate change; genomes are large reservoirs of genetic variability from which organisms can draw their adaptations and thus potentially avoid disappearing.
«Adaptations are characteristics of an organism’s design that allow it to survive and reproduce»
The current climate change process is a great opportunity for research, because we can learn a lot about the genetic mechanisms behind the adaptation of species. These studies can be done by collecting samples of the same species over time or in different geographical locations (colder or warmer ones, for instance) to observe differences in certain genes or genetic markers. With the information from observations in nature, laboratory experiments can be designed to simulate particular climate conditions (usually via temperature changes) and to analyse whether organisms manifest the same reactions related to the same genes or genetic markers. These results can offer information on the genetic basis of adaptive mechanisms, in this case, to climate change. Currently, thanks to so-called «ancient DNA» techniques, we can even analyse genome fragments from fossils of extinct species alongside contemporary species and analyse which of these genome pieces might be key to climate adaptations. We must not forget that the adaptation of an organism does not depend on it having one variant of a gene or another; it depends on the mechanisms regulating these genes. Information regarding gene expression is very valuable because it can help us to foresee the adaptation (or lack thereof) of populations of organisms to the environmental changes derived from global warming.

Genetics can also be very useful from a more practical point of view. If we know which individuals (or populations) present gene variants that grant them more survival opportunities in certain environmental conditions, we can reconstruct some environments that have become extremely degraded due to climate change. This approach has been applied to the reconstruction of several ecosystems in the United States (Wymore et al., 2014). The technique involves the genetic and ecological study of many organisms, from bacteria to plants and animals.
In this text, we describe our research on the genetic mechanisms that allow species to adapt specifically to climate change. We use a type of fruit fly called Drosophila subobscura in which we study chromosomal inversions, i.e., chromosome fragments facing the direction opposite to the usual, as genetic markers.
The species: ‘Drosophila subobscura’
Drosophila subobscura, a fly from the Diptera order, is often used in genetics, especially in evolution studies (Figure 1). Its biogeographical distribution (Krimbas, 1993) covers most of Europe, except the central and northern part of Scandinavia and Iceland. In North Africa it can even be found in the Sahara desert and in the Middle East it is present in Iran. However, the area it occupies in the territories of the former Soviet Union is still largely unknown. It can also be found on Mediterranean islands, as well as in the Azores, Madeira and the Canary Islands. In 1978, its presence was described for the first time in Chile (Mestres, Balanyà, Prevosti, & Serra, 1993) and it quickly spread west of the Andes, from La Serena to Punta Arenas, although its presence in the south of Chile is believed to be seasonal. Such colonisation probably comes from a Mediterranean population, but the exact origin is still unknown. A few years later, in 1982, it was found for the first time on the North-American west coast, from south California to the south of the Canadian state of British Columbia. We must also mention that a small colonising core was detected around the Argentinian city of Mar de Plata. Experts think it could be a secondary colonisation after the main one in Chile. All three regions currently occupied by the species are consistent. All of them have the same climate alternation: an oceanic and Mediterranean climate (up to the semi-arid pre-desert area).
«An inversion is a chromosome fragment facing the opposite direction to normal. It does not represent any problem for the specimens»
One of the main characteristics of D. subobscura is that it only has six polytene chromosomes, called A (X or sexual), J, U, E, O and dot. All of them, with the exception of the dot chromosome, present chromosomal inversions (Figure 2). As we have already mentioned, inversion is the substitution of a chromosome fragment facing the opposite direction to normal. In D. subobscura, the inversions have the names of letters (indicating the chromosome) and a number (identifying each inversion) or the «st» symbol (for standard). A lot of times the same chromosome presents several inversions, forming what is known as an «arrangement». The different inversions forming the arrangements are identified by their numbers separated by the sign «+». Finally, in order to show if the inversions completely or partially overlap, the numbers are underlined (for instance O3+4). The fact that they present inversions does not represent any problem for the specimens because they still have the same genes, only in different order. However, this fact is essential in evolutionary terms because genes in the inversion tend to be inherited together. If the gene combination that contains an inversion is favourable and allows it to adapt better to its environment (survive and reproduce), it will be favoured by natural selection and will increase in frequency over generations. That is why D. subobscura is such a successful model species in evolutionary studies.
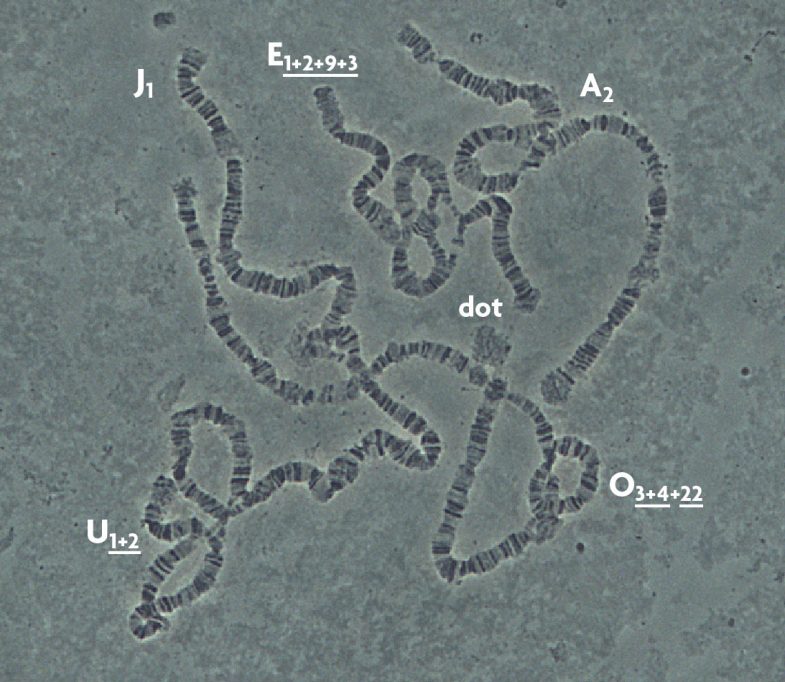
From the 1950s, different European research schools started to analyse inversions in natural populations of D. subobscura. Variation between populations, and even between samples from the same location, was identified. The biggest surprise to scientists was that some of the inversions changed with latitude, that is, they presented what experts call a cline. Thus, some inversions were very abundant in the south of Europe and decreased in frequency in northern samples (such as O3+4). On the other hand, other inversions were not abundant in the south but increased in frequency in northern populations (for example, Ost). There are two possible explanations for this phenomenon: either the involvement of a historical process or because of the adaptive value of the inversions.
In the first scenario an inversion would have appeared at a particular moment (several different mechanisms can generate them, though we will not discuss them in this article) which became very abundant in the population where it appeared. With the natural movement of the Drosophilae, the inversion would have spread passively throughout Europe, thus showing a gradient. Of course, the farther the population spreads from its origin, the less often the inversion would appear. In the second scenario a new inversion that provides an advantage to the specimens that have it would have increased in frequency thanks to natural selection. If the advantage is an adaptation to climate conditions (mainly temperature and other factors linked to it), we would expect its abundance to depend on the position of populations along a north-south axis. For many years, experts discussed which of the two hypotheses explained the cline distribution of inversions in the species (Krimbas & Loukas, 1980). Until two colonised regions were found on the Americas (Chile and the west coast of the United States) which showed latitudinal clines for essentially the same inversions as in Europe, and in the same direction, which strongly supported the adaptive value of inversions (Prevosti et al., 1988).
Climate change and chromosomal inversions
Meteorological measurements imply that the temperature of our planet is increasing; the data indicate the planet has warmed by 0.6 ºC over the last three decades, and by 0.8 ºC compared to the last century (Hansen et al., 2006). Simultaneously, snow and ice seem to have decreased by about 10 %. Likely as a result of these changes, the pattern of rainfall has become more irregular. Many living beings and their ecosystems are affected by this, for instance, by disruption of the pattern of seasonal activity, migrations, or flowering periods.
«If the gene combination that contains an inversion is favourable and allows it to adapt better to its environment, it will be favoured by natural selection»
One of the main researchers who realised the importance of changes in inversion frequencies over time was Antoni Prevosti, and in one of his papers (De Frutos & Prevosti, 1984), he and his colleague describe long-term seasonal changes for the first time. Years later, the last thesis he supervised connected long-term changes in chromosomal inversions with temperature variations (Orengo & Prevosti, 1996), thus once again, highlighting how quickly natural selection acts on D. subobscura. This work was continued by Lluís Serra, who analysed the long-term variation of chromosomal inversions in this species, comparing old data with new for a broad collection of European (thirteen over at least fifteen years) and North and South American (thirteen in a eighteen-twenty year interval) populations. If the warming were global, the frequency of inversions would change according to climate change in both the European and American populations, even several years after the origin of the double colonisation. However, populations analysed in Europe and America usually presented long-term differences both in the type of inversions and in the frequency of «warmth-related» inversions gained over time (Balanyà, Huey, Gilchrist, & Serra, 2009).
Chromosomal inversion studies in Serbia
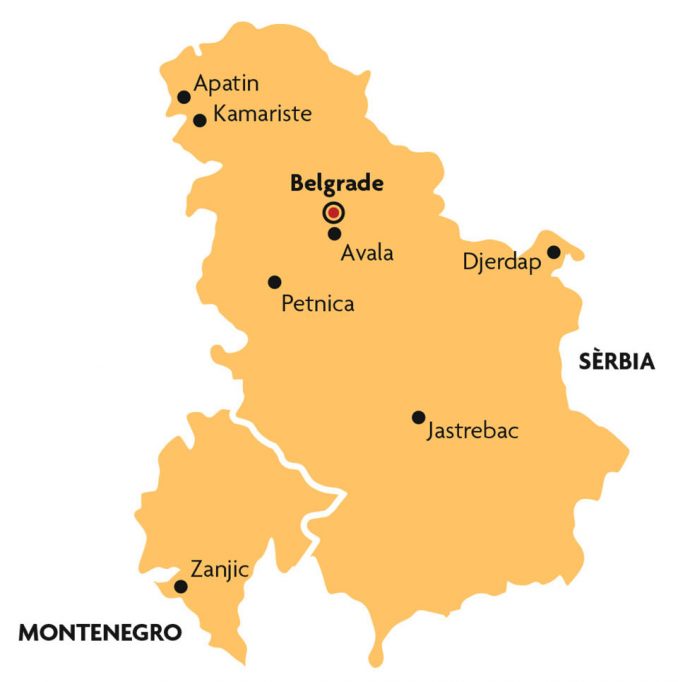
This biogeographical region was important for different reasons. Why was Serbia a particularly important area for study? Most latitude clines historically studied for D. subobscura follow a southwest-northeast axis, starting from the Iberian Peninsula. Furthermore, although it cannot be firmly stated, a number of genetic tests have led scientists to believe that the origin of the American colonisation was a west Mediterranean population. Therefore, Serbia was studied to determine whether or not inversions also changed, and if they followed the predictions for global warming in a different European region where no latitude-cline or time variation studies had yet been carried out. The populations of D. subobscura in the Balkans contain chromosomal inversions that can be considered Mediterranean, but they are very different in some respects (type and frequency of some inversions) to what one can find in the western basin (Krimbas, 1993). Serbia, being part of the Balkans, is considered to have been the refuge for many species during the last glaciation, which makes it attractive in evolutionary terms, because it may have accumulated a lot of genetic variability. Helpfully, inversions have been extensively studied in the Serbian D. subobscura populations thanks to the school founded by Dragoslav Marinkovic, which means a lot of historical information on genetic inversions is available (for a good review, see Krimbas, 1993; Zivanovic, Andjelkovic, & Marinkovic, 2002).

The first study of the series was done with samples from a population in Avala, 18 km away from Belgrade (Figure 3) and 450 m above sea level (Zivanovic & Mestres, 2010a). Collection was carried out during the month of September in 2003, 2004 and 2005. When compared to samples from other Serbian populations, it seemed that «cold-related» inversions (particularly Jst, Ust, Est and Ost) had decreased, while others qualified as «warmth-related» had increased (J1, U1+2, U1+2+6, E1+2+9 and O3+4), with some variation also being observed in consecutive years. This is also to be expected, because environmental conditions during the months before the collection determine the results for chromosomal inversions. We must not forget that many inversions are adaptive, not only with respect to temperature, but to other factors that may or may not be related to it. Seasonal changes were studied in another piece of work, directly related to the aforementioned one (Zivanovic & Mestres, 2010b); in particular, inversions from the same locations (Avala) in a June 2004 sample were compared to another sample from September of the same year, and according the results from other researchers changes in some inversions were also found (on chromosomes J, E and U).

These findings encouraged us to follow this line of research. We chose the population in Apatin, a wetland area on the left side of the Danube, for the next study (Zivanovic & Mestres, 2011). It is a very stable habitat, which is relatively unaffected by human activity, and is ideal for the proposed research project. There was data about the chromosomes analysed in 1994 and new captures were made in 2008 and 2009. As previously mentioned, the first approach to analysing if chromosomal inversions change in line with an influence by climate change is studying whether «warmth-» and «cold-related» inversions increase or decrease, respectively. Nonetheless, to be precise one must also study on-site temperature fluctuations over time. Luckily, we were able to obtain data about maximum, minimum and average temperatures, as well as the rainfall, thanks to the Hydrometeorological Service records obtained from the Republic of Serbia. These showed that the temperatures tended to increase, while the pattern of rainfall was irregular (another potential characteristic of climate change). The composition of the inversions detected also appeared to change over time; the «warmth-related» arrangement U1+8+2 was detected for the first time in 2009, and there was also a significant increase in «warmth-related» inversions U1+2, O3+4 and O3+4+22.
Later we studied the chromosomes of another population, both in terms of monthly variation (short term) and by comparing them to fifteen-year-old samples (long term). The Serbian population chosen was from Petnica (collected in 2010), about 100 km south of Belgrade (Zivanovic, Arenas, & Mestres, 2012), and the specimens were collected in a Carpinus betula forest. This work also aimed to carry out comparative analysis of the Serbian populations we had chromosomal inversion data for (a total of eighteen samples) using multivariate statistical methodology. Interestingly, we detected monthly changes in the chromosome composition of the samples which may be due to natural selection responding to environmental changes, but also to the fact that specimens may move around to look for the temperatures they prefer. As for long term studies, some «warmth-related» inversions which were not observed in the 1995 sample were detected for the first time in 2010 (U1+8+2, E1+2+9+12 and O3+4+8), and an increase in «warmth-related» inversions (A2, J1, U1+2 and O3+4) and decrease in «cold-related» inversions (Ast, Jst, Ust, Est and Ost) was also again observed. One of the highlights of the work is that all of the temperatures studied (maximum, minimum and average) were found to have generally increased, with the most significant change being detected in the minimum temperature.
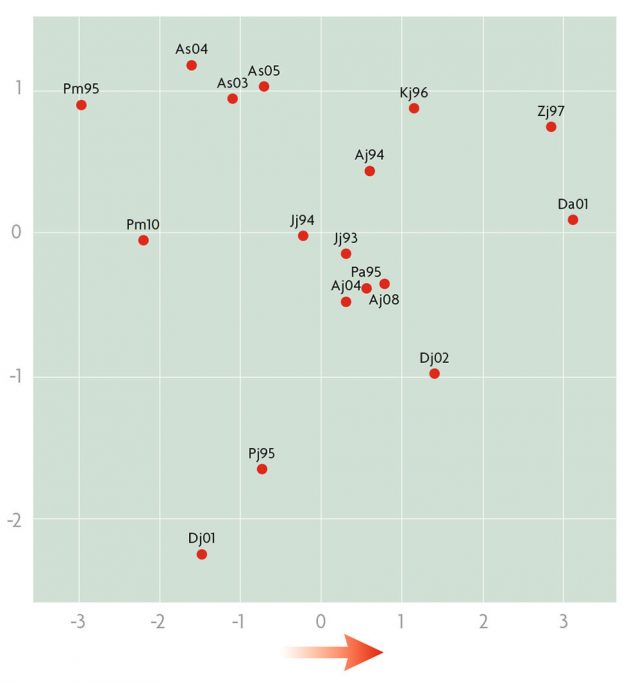
Regarding the study of inversions in the Balkan populations as a whole, our cluster analysis grouped all of the «old» inversions into one group and all of the «recent» inversions into another. Thus, again demonstrating that this genetic marker varies over time. Using inversion data and information about the climate characteristics of the populations, the first and second main components accounted for 67.27 % and 23.17 % of the total variation, respectively (Figure 4). What is interesting is that the first component explains differences between populations which are related to temperature and rainfall, in other words, it changes positively as we move to warmer and drier populations. Therefore, the composition of populations in terms of inversions is a natural selection response to climate change. Moreover, the variation moves in the direction we would expect if it were due to the effect of global warming on our planet.
This field of study is fascinating and our intention is to continue these experiments and analyses, for example, by observing the effect of inversion combinations from the same homologous pair (karyotypes) or the interactions between chromosome inversions from different pairs. Indeed, our preliminary studies seem to indicate that inversions present more adaptive value than karyotypes (Zivanovic, Arenas, & Mestres, 2014). In short, chromosomal inversions are a good marker of climate change and, in addition, they allow us to study how species adapt to it.
REFERENCES
Balanyà, J., Huey, R. B., Gilchrist, G. W., & Serra, L. (2009). The chromosomal polymorphism of Drosophila subobscura: A microevolutionary weapon to monitor global change. Heredity, 103,
364–367. doi: 10.1038/hdy.2009.86
De Frutos, R., & Prevosti, A. (1984). Temporal changes of chromosomal polymorphism in natural populations of Drosophila subobscura. Genetica, 63, 181–187. doi: 10.1007/BF00128411
Hansen, J., Sato, M., Ruedy, R., Lo, K., Lea, D. W., & Medina-Elizade, M. (2006). Global temperature change. Proceedings National Academy of Sciences, 103, 14288–14293. doi: 10.1073/pnas.0606291103
Krimbas, C. B. (1993). Drosophila subobscura: Biology, genetics and inversion polymorphism. Hamburg: Verlag Dr Kovac.
Krimbas, C. B., & Loukas, M. (1980). The inversion polymorphism of Drosophila subobscura. Evolutionary Biology, 12, 163–234.
Mestres, F., Balanyà, J., Prevosti, A., & Serra, L. (1993). Genética evolutiva de la especie colonizadora Drosophila subobscura. Mundo Científico, 13, 408–416.
Orengo, D. J., & Prevosti, A. (1996). Temporal changes in chromosomal polymorphism of Drosophila subobscura related to climatic changes. Evolution, 50, 1346–1350. doi: 10.2307/2410676
Prevosti, A., Ribo, G., Serra, L., Aguade, M., Balaña, J., Monlcus, M., & Mestres, F. (1988). Colonization of America by Drosophila subobscura: Experiment in natural populations that supports the adaptive role of chromosomal-inversion polymorphism. Proceedings National Academy of Sciences, 85, 5597–5600. doi: 10.1073/pnas.85.15.5597
Wymore, A. S., Bothwell, H. M., Compson, Z. G., Lamit, L. J., Walker, F. M., Woolbright S. A., & Witham T. G. (2014). Community genetics applications for forest biodiversity and policy: Planning for the future. In T. Fenning (Ed.), Challenges and opportunities for the world’s forest in the 21st century (pp. 707–726). London: Springer.
Zivanovic, G., Andjelkovic, M., & Marinkovic, D. (2002). Chromosomal inversion polymorphism of Drosophila subobscura from south-eastern part of Europe. Journal of Zoological Systematics and Evolutionary Research, 40(4), 201–204. doi: 10.1046/j.1439-0469.2002.00189.x
Zivanovic, G., & Mestres, F. (2010a). Viabilities of Drosophila subobscura homo- and heterokaryotypes at optimal and stress temperatures. I. Analysis over several years. Hereditas, 147, 70–81. doi: 10.1111/j.1601-5223.2009.02163.x
Zivanovic, G. & Mestres, F. (2010b). Viabilities of Drosophila subobscura homo- and heterokaryotypes at optimal and stress temperatures. II. Seasonal component analysis. Hereditas, 147, 82–89. doi: 10.1111/j.1601-5223.2010.02164.x
Zivanovic, G., & Mestres, F. (2011). Changes in chromosomal polymorphism and global warming: The case of Drosophila subobscura from Apatin (Serbia). Genetics and Molecular Biology, 34, 489–495. doi: 10.1590/S1415-47572011000300020
Zivanovic, G., Arenas, C., & Mestres, F. (2012). Short- and long-term changes in chromosomal inversion polymorphism and global warming: Drosophila subobscura from the Balkans. Israel Journal of Ecology and Evolution, 58, 289–311. doi: 10.1560/IJEE.58.4.289
Zivanovic, G., Arenas, C., & Mestres, F. (2014). Inversion polymorphism in two Serbian natural populations of Drosophila subobscura: Analysis of long-term changes. Russian Journal of Genetics, 50, 638–644. doi: 10.7868/S0016675814060150




