

Should I Leave or Should I Stay?
Understanding dispersal behaviour

Many factors influence dispersal decisions, and studying these factors can reveal the causes and determinants of dispersal. Animals leave their birthplace to solve three fundamental problems: habitat deterioration, competition, and risk of inbreeding. However, animals only disperse once they are physically fit enough to reach and settle in the new location successfully. The dispersal process has three phases: departure from the natal area; transition (displacement); and settlement in the new habitat. Animals face different decisions in each of these phases.
Keywords: habitat quality, competition, inbreeding, dispersal, fitness, gene flow.
«Dispersal occurs in virtually all species, and is probably one of the most important life history traits involved in the evolution of species»
Dispersal is one of the most widely studied concepts in ecology and evolutionary biology, but we still have much to learn. Dispersal is the movement of individuals, or of specialised structures for dispersal (diaspora, such as seeds or pollen) to a new territory where reproduction takes place at least once. When we speak of displacement in animals, we should avoid confusion between dispersal and migration. Migration is characterised by a periodic displacement of many individuals simultaneously, and does not always result in gene flow (exchange of gene variants) between groups or populations. Dispersal, however, involves few individuals, and often results in gene flow as individuals join a new population within which they reproduce. For example, swallows breed throughout Spain, nesting in barns, stables and farmhouses, and live in more or less numerous groups. Then, in late summer they migrate to sub-Saharan Africa, where winter conditions are milder. When early spring arrives, they return to their breeding areas, but some individuals may disperse, in other words they move and settle on another farm or farmhouse located a few metres or kilometres away from the previous one, where they may breed with new potential partners. In this case, we can say that these individuals have made a seasonal migration and then, on returning to breed, have dispersed. Therefore, dispersal involves displacement and gene flow. These features may have profound ecological and evolutionary implications for the species under consideration. In fact, dispersal occurs in virtually all species, and is probably one of the most important life history traits involved in the evolution of species.
«Individuals must first decide whether to disperse or not, then when to start dispersing, and finally when to stop and settle»
Unlike the study of dispersal in plants, approaches to studying animal dispersal have to consider their ability to move. In animals it is the individuals themselves that move with bodily effort, while in plants displacement is experienced by diasporas, namely, propagules (fruits, seeds or pollen), and is mediated by environmental factors (e.g., wind or water) or animals. Thus, in plants a single dispersal episode is common, whereas animals may disperse before each breeding event. While the ability to move is a fundamental feature differentiating dispersal in plants and animals, it is interesting that the pattern showing dispersal distances in both groups of organisms reflects the same feature: the predominance of short distances over long distances (Fig. 1). Somehow, both groups benefit from dispersal without the need for large displacements, which generally imply greater costs.
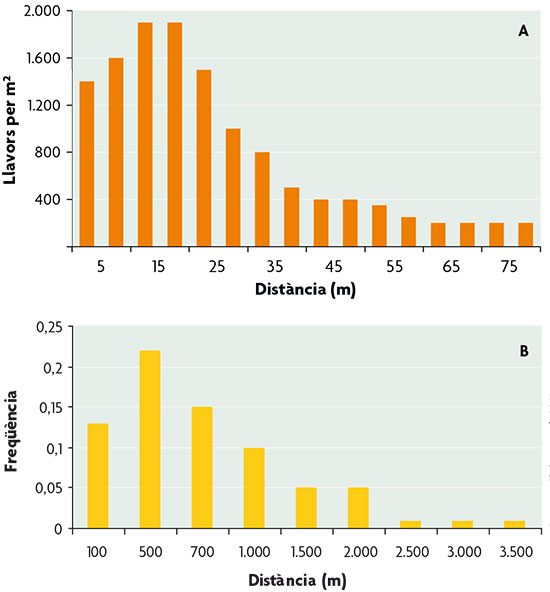
Figure 1 Distribution of dispersal distances in an angiosperm plant (A: sorrel, Tachigali versicolor) and in a passerine bird (B: Great Tit, Parus major). / Basat en Clobert et al., 2001
Linked to the ability to displace and move actively, another key factor in animal dispersal is the ability to «decide». Firstly, individuals must decide whether to disperse or not, secondly, when to start dispersing, and finally when to stop and settle. Similar issues exist in plant dispersal, but are at the mercy of the mediating factors. The consequences of dispersal are none other than the consequences of how organisms solve these issues, and the effects on their fitness, in other words, survival and reproduction, are ultimately the evolutionary forces of dispersal.
The dispersal process comprises three phases: departure from the home area, transition (displacement) and settlement in the new habitat. Different decisions must be taken at each of these phases. Which members of a population decide to disperse? Why? When does dispersal begin? When does it stop? Animal dispersal studies explore the factors influencing each of these decisions. A good approach to predicting the result of these decisions is to consider the costs versus the benefits of potential decisions. Disperse or stay put? Multiple factors influence the decision to leave the natal area, and in some cases these factors act in opposing ways. For example, the benefits gained by reaching new territories act in favour of dispersal, while energy expenditure and mortality risk during displacement act against dispersal.
«Often, the individuals who disperse are not a random sample, but are more efficient at moving and colonising new territories»
By considering the dilemmas that individuals face, biologists can get a picture of the big issues that characterise dispersal: the ultimate causes (its purpose) and immediate causes (what factors determine it). It is not easy to study dispersal, because it requires objective measurements of displacement and reproductive success of the individuals dispersing into new territories. Thanks to demographic and genetic methods, we have data for an ever increasing number of species. The scientific study of dispersal in different animal species has provided evidence for interesting associations between dispersal-related variables, which can hardly be considered purely random, as discussed below.
Who will leave? Characteristics of individuals that disperse
Individuals forming a population differ in their morphological, physiological and/or behavioural traits (i.e., they have different phenotypes), or usually show great intraspecific variability. Often, the individuals who disperse are not a random sample, but are more efficient at moving, colonising new territories and integrating into a new population. In other words, these individual phenotypes are more likely to successfully complete the act of dispersal.
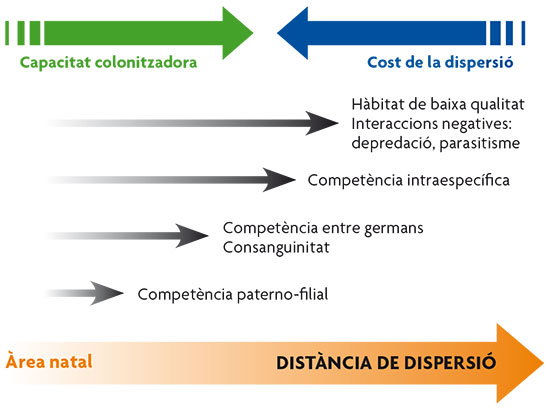
Figure 2. Diagram showing different dispersal distances necessary to escape the negative characteristics of the natal area. / Basat en Clobert et al., 2001
In some species, the individuals that disperse are morphologically different from those that do not. For example, some insects like greenflies (aphids) have winged and wingless members and dispersal is undertaken by a higher proportion of winged members, which can achieve greater dispersal. In other cases, individuals that disperse display different behavioural traits. For example, in species living in social groups, dispersal to a new habitat involves having to join a new social group, so a willingness to live with individuals outside their own group may be beneficial. This is the case of the naked mole-rat, a mammal living in social groups. Studies have found behavioural differences between the individuals that disperse and those that do not. In experiments, mole-rats were given the choice to mate with individuals of their own group or from another group, and those that dispersed showed a preference for the mate from another group (O’Riain et al., 1996). This behaviour would undoubtedly facilitate its integration in any group in a new habitat.
What is gained by leaving? Benefits and costs of leaving the natal land
As mentioned above, the decision whether or not to leave one’s birthplace involves a cost-benefit balance. The costs of dispersal are numerous and involve more than just the energy cost of displacement and predation risk. For instance, costs also arise from a lack of knowledge of the new habitat, involving the need to find new food resources or new social groups whose cooperation is required for certain activities. There is also the cost of losing what we call local adaptation, which refers to those subtle adjustments developed in response to the specific environmental conditions of the habitat of origin, which are lost in the new habitat. This is the case of ectoparasites such as fleas, which may have developed a high level of adaptation to the specific conditions of their host, for instance the body temperature of a dog, and thus dispersal to a new host may entail the cost of adapting to new local conditions. In general, the cost of the dispersal increases with distance, as reflected in Figure 2.
Moreover, dispersal is considered to serve three main functions (ultimate causes): improve habitat quality, avoid intraspecific competition and prevent inbreeding (Clobert et al, 2001). Below, we will take a look at some examples of evidence for each of these factors underlying dispersal. However, we can expect differences in dispersal behaviour depending on which of these causes exerts greatest pressure, for example, in the distances covered (Figure 2). So, moving a short distance, such as to the adjacent territory, can solve the problem of competing with parents in territorial animals. Greater distances would be required to beat the competition between siblings and the risk of inbreeding, as this often implies changing social group. The longest distances individuals travel are those undertaken to colonise better quality lands.
In Search of El Dorado: when the aim is to improve habitat quality
Interesting evidence demonstrates there is a relationship between dispersal and habitat quality. Indeed experimental manipulations of quality-related components, such as the presence of species that interact negatively or the availability of resources such as food, shelter or breeding places. For example, in a population of aphids, a decline in habitat quality due to an increase in one of their main predators, ladybugs, leads to the birth of a higher proportion of winged individuals in the next generation (Weisser et al., 1999). Conversely, the elimination of parasites in bird nests, which clearly improves the breeding habitat, delays the onset of the dispersal of chicks.
In terms of resources, increased food availability can lead to a decrease in dispersal. This is the case of gulls, which currently cut their dispersal distances, due to a rise in the number of landfills in coastal areas. Similarly, the practice of discarding fish – common to fishing boats – has taught seabirds such as shearwaters of this daily food supply, and this has diminished their dispersal to areas without fishing. The availability of shelters is also a determining factor in the occupation of a territory, in reptiles for instance; thus, placing logs to create artificial shelters can significantly increase the arrival of reptiles in an area. The same applies to the availability of breeding sites, which may encourage birds to disperse to places where the availability of these sites has been experimentally increased by placing nesting boxes.
When the family is the problem
Some experimental studies show how competition between individuals of the same species can influence dispersal. For example, in numerous insect species, dispersal increases when the density of individuals is increased artificially, while dispersal diminishes with an experimental increase in the amount of food available.
A great deal of evidence comes from one particular type of competition: intraspecific competition among related individuals, in other words, between siblings, and between parents and offspring. For example, some parents force the departure of some of their offspring to prevent competition between siblings, even though this forced dispersal may lead to mortality. This has been observed in some birds, such as jays, where fitness advantages tend to be greater for the parents (ensuring the survival of the fraction that stays) than for the dispersed individual. Other evidence that dispersal may respond to competition between siblings is to be found in species exhibiting interactions between dominant and subordinate siblings, with the subordinate being forced to leave. Indirect evidence of the importance of sibling competition can be observed in some mammals, such as foxes, where smaller litter sizes coincide with a lower tendency to disperse.
Competition between parents and children usually occurs in species that are heavily dependent on a territory. In these species, offspring may attempt to dominate territory early on, creating conflict among siblings and with parents. In some cases, like the ground squirrel, parents only force the more precocious young to disperse, as they are more likely to attain territory than the late-developers. In other cases, such as the song sparrow, parents aggressively drive hatchlings out of the territory. In a third group of cases, like the red squirrel, the parents are the ones to leave, and do so rather than entering into competition with their offspring. There is also indirect evidence that competition among relatives favours dispersal in that cooperation between them decreases dispersal, as in lions and some primate species.

In some species the individuals that disperse are morphologically different from those that do not. For example, there are winged and wingless aphids, and a higher proportion of winged individuals disperse, which are those that can travel further. / David Martínez Torres
When incest is the danger: avoiding risks of inbreeding
In many species, dispersal is gender biased, which seems a perfect way to avoid inbreeding. Interestingly, males are more likely to disperse in some species and females in others. The explanation in each case can be found in terms of the costs and benefits of dispersal for each sex, so dispersal of one particular gender decreases the probability of mating with relatives while providing specific benefits to the gender that leaves. These benefits are influenced by the mating system and social structure characterising the species. When monogamy is the mating system (a male mates with a single female) female dispersal is more prevalent, while in polygyny (a male mates with several females) dispersal of males is more common. Regarding social structure, female dispersal increases when social structure is based on the defence of resources, whereas male dispersal is favoured when the social structure is rooted in the defence of females by males.
«Dispersal serves three main functions: To improve habitat quality, avoid intraspecific competition and prevent inbreeding»
Birds and mammals are the groups that provide most evidence for the importance of these factors. In birds, males compete for territory, so they benefit more from being familiar with it than females do. Therefore, fewer males disperse than females. By contrast, females choose the male according to his territory (or his nest-building ability) and, therefore, dispersal allows them to choose between a larger number of territories or nests offered. So in this case, females disperse more than males. Conversely, in mammals, it is the females that compete for territory and thus they benefit more than males by familiarity with it and so disperse less than males (Cockburn et al., 1985). They, however, choose territories where females are present, and thus benefit from dispersal as it enables them to access a greater number of territories with females. Consequently, males disperse more than females.
Disperse, yes, but when should i leave?
Dispersal is determined by three groups of factors (proximate causes): environmental factors, internal factors of the individuals that disperse, and genetic factors.
Environmental conditions are one cause of dispersal. However, animals do not always respond directly to environmental conditions, but in some cases it is an indirect response, mediated by a preceding effect on the individual’s physical condition. Generally speaking, a direct dispersal response is brought on by sudden or unpredictable changes in environmental conditions such as invasion by natural enemies, limited availability of mates or extreme weather events, whereas an indirect dispersal response is triggered by gradual and predictable changes in environmental conditions, such as seasonality or resource availability.
Two types of internal factors trigger dispersal: an individual’s physical condition and the levels of certain hormones in the blood. A robust physical condition seems to be a pre-requisite to undertaking dispersal. Evidence indicates that some individuals delay departure from the natal area until they have stored adequate energy reserves. Regarding hormones, high levels of those related to displaying aggression, such as testosterone, and to locomotor activity and response to stressful situations, such as corticosterone, are also required to start dispersal. There is interesting evidence in mammals, birds and reptiles for the role of these hormones in dispersal behaviour (Belliure and Clobert, 2004). For example, embryos of mammals like mice exposed to testosterone during pregnancy are more prone to disperse. Likewise, in birds, the level of testosterone in eggs influences chick dispersibility, which increases in parallel to testosterone levels. In young lizards, corticosterone levels are related to locomotion capacity and dispersal propensity.

Greater food availability can lead to a decline in dispersion. This happens in seagulls which now travel shorter dispersal distances as they can scavenge from landfills in coastal areas. / Grand Parc de Burdeos
Moreover, if we consider the possible genetic basis for dispersal we should wonder whether there is genetic variation in the characteristics related to dispersal, and if the traits involved in dispersal are always expressed together, that is, if they are genetically correlated. Evidence points to the existence, at least in certain animal species, of components of genetic determination. For example, in aphids genetic variation exists between winged and wingless individuals. And in some species of Orthoptera insects, like crickets, there is evidence for genetic correlation between three dispersal-related characteristics: a predisposition to fly, wing shape and powerful muscle physiology. These three traits, involved in the dispersal of crickets, are always expressed together.
In short, to stay or to leave one’s birthplace is one of the key decisions most organisms must take, including ourselves as members of an animal species that also provides ample evidence of what we have discussed. Either option may be apt within the context of different environmental scenarios, but a suitable response is key to a successful life. Thus the factors influencing dispersal are powerful evolutionary forces and the dispersal behavioural trait has profound consequences for species. Further study of animal dispersal will undoubtedly continue to provide remarkable evidence for the power of these evolutionary forces in shaping such a complex trait.
Belliure, J. i J. Clobert, 2004. «Behavioral Sensitivity to Corticosterone in Juveniles of The Wall Lizard». Physiology and Behavior, 81: 121-127. DOI: <10.1016/j.physbeh.2004.01.008>.
Clobert, J.; Danchin, E.; Dhondt, A. A. i J. D. Nichols (eds.), 2001. Dispersal. Oxford University press. Oxford.
Cockburn, A.; Scott, M. P. i D. J. Scotts, 1985. «Inbreeding Avoidance and Male-Biased Natal Dispersal in Antechinus spp. (Marsupialia: Dasyuridae)». Animal Behaviour, 33(3): 908-915. DOI: <10.1016/S0003-3472(85)80025-7>.
O’Riain, M. J.; Jarvis, J. U. M. i C. G. Faulkes, 1996. «A Dispersive Morph in The Naked Mole-rat». Nature, 380: 619-621. DOI: <10.1038/380619a0>.
Weisser, W. W; Braendle, C. i N. Minoretti, 1999. «Predator-induced Morphological Shift in The Pea Aphid». Proceedings of the Royal Society London B, 266: 1175-1181. DOI: <10.1098/rspb.1999.0760>.




