

Out of Africa
An alternative scenario for the first human dispersal in Eurasia
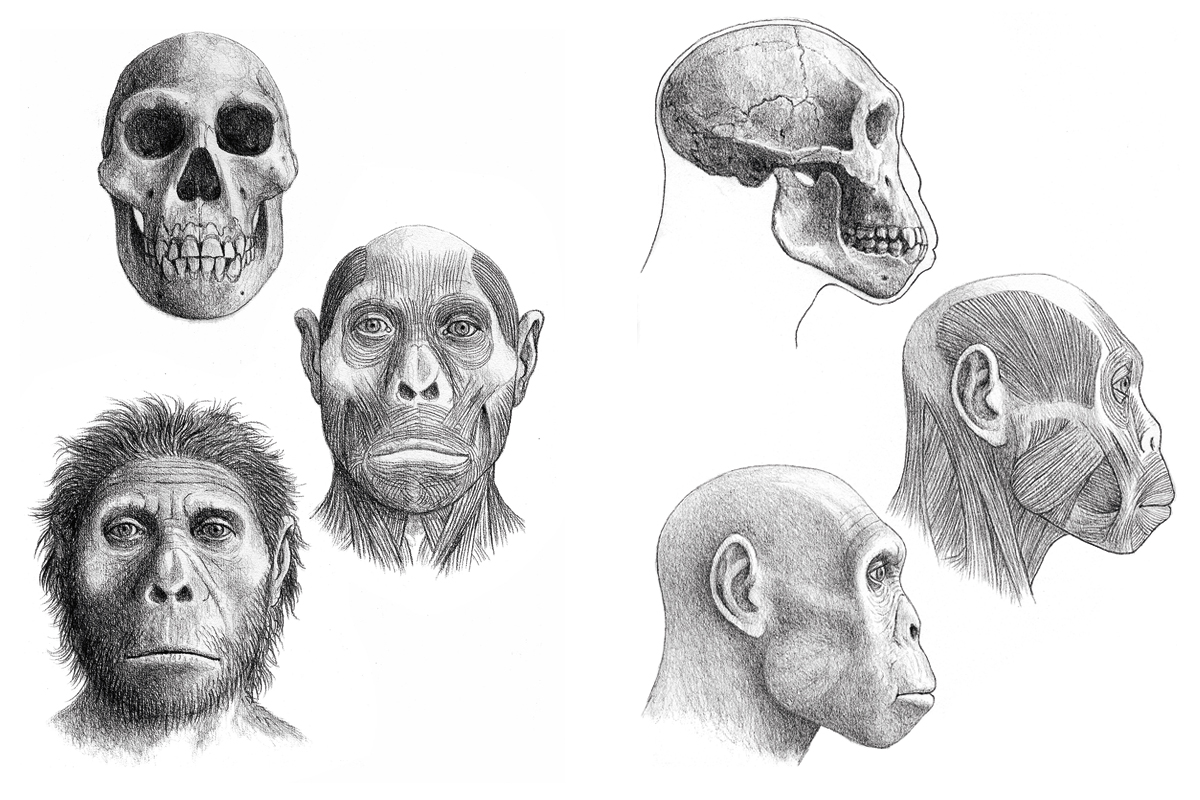
Recent paleoanthropological evidence from the early Pleistocene site of Dmanisi in Georgia has revealed that the first hominins out of Africa were more archaic than the coeval African and Asian Homo erectus. More evidence suggests that these archaic hominins were forest dwellers rather than savannah inhabitants. Between 1.8 and 1.6 million years ago a climate crisis caused a new spread of savannah and arid zones across large parts of Africa. As a consequence, early Homo populations splitted, with some populations becoming adapted to the new ecological conditions and others following woodland areas in their regression.
Keywords: out of Africa, Dmanisi, early Homo, sabertooth cats, Early Pleistocene, paleoclimatology.
Introduction
Our ideas about how and when the first human dispersal out of Africa took place have changed drastically since the 1990s as a result of the discoveries at the Dmanisi site in Georgia, which have proven the presence of extremely archaic hominids in the southern Caucasus almost two million years ago. In particular, the discovery of the quite complete D2700 skull forced a reassessment of the scenario which placed African Homo erectus and its anatomical and cultural novelties as the cause for such «out of Africa» (Vekua et al., 2002). Recent data about skull D4500 also confirms that point of view (Lordkipanidze et al., 2013). As D2700, D4500 present features that are clearly more archaic than African Homo erectus, and its cranial capacity is also smaller than the average for this African species. The Dmanisi discoveries thus refuted the idea that Homo erectus innovative features were what propitiated the first exit from Africa (Lordkipanidze et al., 2007). Alternative scenarios were also refuted by Dmanisi findings (Agustí & Lordkipanidze, 2011). Carbonell et al. (1999), for example, proposed a cultural scenario in which the upsurge of Mode II or Acheulian, with the technological innovation that led to the creation of bifaces, triggered the movement out of Africa of the more primitive populations that still used Mode I or Oldowan tools. However, the first evidence of Mode II tools (in Konso-Gardula, Ethiopia) is dated at around 1.4 million years ago, while the Dmanisi hominids date back to roughly 1.8 million years ago.
«The Dmanisi discoveries thus refuted the idea that ‘Homo erectus’ innovative features were what propitiated the first exit from Africa»
Departure from Africa was thus almost 400,000 years before the expansion of Mode II and hence cannot be linked to this cultural overhaul. At the opposite extreme, Martínez-Navarro and Palmqvist (1995; see also Arribas & Palmqvist, 1999) have proposed an ecogeographic scenario, according to which hominids merely followed the expansion into Eurasia of some of the fauna which, like them, was part of the African ecosystems. This would have been the case for the hippopotamus Hippopotamus antiquus, the hyaena Pachycrocuta brevirostris, the sabertooth cat Megantereon whitei, the giant baboon Theropithecus oswaldi, and a range of species with a clear African origin that have been found at the Ubediya site in Israel, such as the large bovine Pelorovis oldowayensis, the antelope Oryx sp., the large boar Kolpochoerus oldowayensis, the wild ass Equus tabeti, and others (Tchernov, 1992). According to this scenario, predators and hominids followed the northward migration of the large herbivores that were a common part of their diet, finally arriving in the lands that spread beyond the African continent.
However, these different events scarcely constituted anything like a «migratory wave». On the contrary, the majority were isolated, independent cases of migration. The spread of the large hyena P. brevirostris, for example, took place a little less than two million years ago, while the hippopotamus H. antiquus followed almost 400,000 years later. The first evidence of the baboon Theropithecus in Spain dates back almost a million years, 600,000 years after the arrival of H. antiquus. It is thus difficult to acknowledge the existence of a single «wave of migration» that could have transported part of the early Pleistocene savannah ecosystems from Africa to southern Europe, hominids and all. But it is also difficult to detect the African imprint of such migration in Dmanisi (Agustí & Lordkipanidze, 2011; Tappen, Lordkipanidze, Bukshianidze, & Ferring, 2007).
«There is broad consensus that it would have been difficult for these early representatives of our genus to have been active hunters»
The presence of African species in Dmanisi (obviously apart from hominins) has been proclaimed by the proponents of the migratory wave, who mention the presence of the sabretooth cat M. whitei, the giraffoid Paleotragus sp., and the giant ostrich Struthio dmanisiensis. However, the sabre-tooth Megantereon from Dmanisi has been attributed by Vekua et al. (2002) to the European species M. cultridens. The giraffoid Paleotragus sp. is more a Pliocene relict than a giraffe of African origin. We know that representatives of this genus appear to have persisted south of the Himalayas during the Pliocene, after their extinction in Europe at the end of the Miocene. In contrast, a glance at the rest of the Dmanisi fauna reveals an overwhelming majority of common elements to Europe’s early Pleistocene fauna: archaic mammoths (Mammuthus meridionalis), horses (Equus stenonis), rhinoceros (Stephanorhinus etruscus), deer (Eucladoceros senezensis, Cervus perrieri, Pseudodama nesti), gazelles (Gazella borbonica), ovibovines (Soergelia sp.), sabre-tooths (Homotherium latidens) and dogs (Canis etruscus). Even the hyaenas from Dmanisi belong to be typical European Pliocene form, Pachycrocuta perrieri, and not the large hyaena P. brevirostris.

Early ‘Homo’ as a scavenger
Having discarded the scenarios that link the first movement out of Africa to Homo erectus evolutionary innovations, cultural factors, or migratory waves by part of the African ecosystem, the question about the cause of this movement still remains to be answered. To provide an alternative explanation to the first human movement out Africa, which would be consistent with the recent Dmanisi discoveries, it may be more useful to focus on the original lifestyle and habitat occupied by these archaic populations of our genus at the time when the migration took place.

There is broad consensus that it would have been difficult for these early representatives of our genus to have been active hunters. In contrast, scavenging may have supplied the protein input required to maintain an energy-expensive organ like the brain. However, scavenging in an open savannah like the one that covered a large part of Africa during the Pleistocene was by no means easy. Many observations have shown that, after a hunting episode, the carcasses are quickly detected by a cohort of scavengers, particularly vultures and hyenas. In this open savannah context, the first hominids which exploited the scavenger’s ecological niche would have found it quite hard to get to the carrion. The ability to act as a kleptoparasite, i.e., chasing away a large felid to get at its prey, would obviously have been close to the limits of its possibilities. But even getting to the predator’s leftovers would have required active scavenger behaviour by these hominids, i.e., they would have had to openly confront other scavengers such as hyenas, jackals and wild dogs, competing with them to exploit the carcass. They could only behave as passive scavengers, i.e., scavengers that do not have to compete with other carnivores of a similar or larger size at certain times of the year (e.g., at the end of the dry season, when many large herbivores die from hunger and not as victims of large predators).
This panorama would change completely, however, if, instead of trying to scavenge in the open savannah, we move to forest zones that often have clearings near watercourses and lakes, where the situation is quite different. To begin with, carcasses are detected by potential scavengers much later due to the lack of vultures, veritable living signposts. Several observations (Blumenschine, 1988; Blumenschine & Marean, 1993; Domínguez-Rodrigo, 2001) point out that a carcass can remain untouched for up to one or two days without any carnivore showing interest in it. A scavenger discovering such a resource has much more time and calm for defleshing. Competition for carrion is thus much lower than in the open savannah.
It is thus clear that at the beginning of the Pleistocene, forest habitats provided many more scavenging opportunities for hominids. In dangerous situations, the archaic Homo habilis, with an anatomy that was still reminiscent of Australopithecus, could quickly climb trees and even store away the retrieved material, just as leopards do today. In fact, the huge variability shown by the Dmanisi paleodeme (Lordkipanidze et al., 2013) is only found in living antropomorphs inhabiting forest environments. Forests are, moreover, much safer places than savannas, where hominins are on open ground and can be seen from afar by a predator.
The role of sabertooth cats
Early Pleistocene forest biotopes also had other inhabitants such as the sabertoothed felids Megantereon and Homotherium. In fact, studies of the motor skeletons of these saber-toothed have revealed an anatomy that was better prepared for life in forests and closed habitats than on the savannah and open spaces, where large modern types of felids cope much better. This is particularly obvious in the case of Megantereon, a relatively small and robust species with feet that allowed it to climb trees (Agustí & Antón, 2002; Marean, 1989). While Megantereon probably inhabited dense forests, the Homotherium anatomy was more appropriate for running, which allowed it to inhabit arboreal savannahs and open forests. Homotherium and Megantereon also had a vertical distribution of their common habitat: while the former was more comfortable on the ground, the latter lived in the tree-tops.
«At the beginning of the Pleistocene, forest habitats provided many more scavenging opportunities for hominids»
Although it may seem strange, the presence of Megantereon and Homotherium in the same forests and arboreal savannah occupied by hominids two million years ago was more an advantage than a disadvantage for the latter, in spite of the occasional inevitable deaths from predation. Sabertooths were able to supply hominids with a lot more carrion than the modern savannah felids such as leopards (Marean, 1989; Marean & Erhardt, 1995). The sabertooth dentition was well adapted to bringing down and tearing apart large prey. Its canines were veritable knives, capable of cutting even the toughest hide. By the same token, however, these efficient teeth were poorly adapted to removing meat attached to bone because, unlike today’s felines, their large canines could not be used for this purpose, and they can be attributed with even less ability to grind and ingest the bones themselves.

Sabertooths would have therefore been much less efficient than modern felines at finishing off the carcasses of their prey, giving hominids a double advantage. On the one hand, they would have left considerable amounts of unconsumed meat, while on the other hand, these sabertooths would have left their prey quite quickly at the mercy of potential scavengers. It is not hard to imagine agile Homo habilis sitting in trees, waiting for the right moment to leap onto the prey with their stone cores and flakes following the departure of a satiated Homotherium or Megantereon. Eventually, hominins also had access to the prey left by the Megantereon on branches, like modern-day leopards. Forests and arboreal savannahs near lakes and watercourses were thus a much more appropriate environment for scavenging than the open savannah, and they are quite likely to have been the original environment where the first representatives of our genus developed. This environmental context is most probably related with the causes that led hominins to move out of Africa.
Savannah expansion in the Early Pleistocene
Between 1.8 and 1.6 million years ago a climate crisis caused a new spread of arid zones across large parts of Africa, resulting in the further retreat of hominins from the tree-covered areas which had been the favourite habitat for sabertooths. This change proved to be lethal for Megantereon and Homotherium: unable to compete with modern felines on the open savannah, these two large carnivores became extinct in sub-Saharan Africa about 1.5 million years ago. Meanwhile, Homo habilis was forced to enter the savannah and behave as an active scavenger, forced to confront large carnivores and compete fiercely with other potential scavengers for the carcasses. Under these circumstances, it is quite likely that active scavenging was replaced by direct hunting, which may have ultimately been much more effective through the obligation of confrontation with other predators. This is a key point that has been proposed to explain the transformation of Homo habilis, a passive scavenger and forest dweller, into Homo erectus, an active scavenger and potential hunter who, unlike the former species, was perfectly well adapted for life on the open savannah (Marean, 1989). This is also the point where the issue of the first movement out of Africa reappears.
«Sabertooths were able to supply hominids with a lot more carrion than the modern savannah felids»
It is quite possible that, while some Homo habilis populations were effectively forced to enter the savannah and adapt to the new conditions, others may have tried to follow the retreating forest habitats. The spread of arid and even desert zones in the Sahara belt probably caused a split. South of this belt, some populations found refuge in the tropical parts of Africa, although they would have had to compete there with other primates which were better adapted to life in the trees. North of this belt, however, beyond the bounds of the continent, these forest hominins may have discovered a similar biotope to the one that was shrinking at that time across a large part of Africa, still inhabited by sabertooths and favourable to passive scavenging. Such a refugium could have been the southern Caucasus.
Today, the Caucasus area still has exceptional features. South of this large mountain chain is one of the last refuges of subtropical vegetation that covered a large part of Europe during the Miocene and Pliocene. This is a consequence of the special climate conditions in this area, fringed by the large water bodies of the Caspian and Black Seas which supply the moisture required by this type of flora. At the same time, the imposing wall of the Great Caucasus is an effective barrier to the cold, dry winds from the north that blow across the Russian steppes. The conditions here were even more favourable at the early Pleistocene, when an arm of shallow water north of the Caucasus connected the Black Sea and the Caspian Sea, which stretched much further west to form the deep Kura Gulf. At that time, the Great Caucasus was a broad peninsula connected to the Middle East through the Dmanisi region.

The study of fossil pollen at a number of sites has shown the persistence during this period of many subtropical plants such as Tsuga, Marsh Cypress, Engelhardtia, and Alcanfor trees, as well as other temperate species including spruce, oak, beech, pine, and elm. We know that this type of mixed forest persisted in the Caucasus until a relatively late date, in the middle Pleistocene, roughly 400,000 years ago (Shatilova et al., 2011). At the time, there was a clear contrast between seasons, with hot summers and high rainfall. In this context, the Caucasus was a perfect haven for these archaic forest hominids whose habitat was gradually shrinking in Africa. Northward dispersal seems to be the natural route that would have permitted the maintenance of the original living conditions qualities of these early Homo populations. The associated fauna types, predominantly large herbivores and sabertoothed felids, also matched the African habitats where these hominins developed their initial scavenging skills.
In order to reach the southern Caucasus zone, however, the Dmanisi hominids would have had to cross the Middle East, the so-called «Levant Corridor». Today, this area, an extension of the Sahara desert that runs south through the Negev and Syria deserts, has an extremely arid climate. Rainfall above the 100 mm annual average in the form of summer storms is restricted to the northern part of Israel and the Taurus and Zagros mountain ranges, while rainfall in the South is barely 40 mm a year. It is thought that during the climate crisis 1.8 million years ago, with the Sahara in the drier parts of Africa spreading their boundaries at the expense of the already reduced forests, conditions along the Levant Corridor would not have been much better than at present.
«The Caucasus was a perfect haven for these archaic forest hominids whose habitat was gradually shrinking in Africa»
However, this apparent barrier could have been accessible thanks to the particular climatic circumstances of the zone. Fossil pollen analysis in Israel (Dennell, 2009; Horowitz, 1989) revealed that, as in the Caucasus, forests were also predominant in the Middle East during the early Pleistocene. This apparent climatic contradiction is explained by arguing that while the spread of the ice caps usually mean increased aridness in the inter-tropical areas and the spread of arid zones, in the Levant Corridor the effects are exactly the opposite, resulting in increased rainfall and the development of temperate forests (Horowitz, 1989). This is because during the glacial phases, the northern hemisphere climate belts move further south from their current position. In this situation, the cold but moist Atlantic winds and the polar fronts spread across the entire European plain to the Middle East which paradoxically experiences higher rainfall and a much moister climate than in the interglacial periods.
Conclusions
In conclusion, the evidence here presented points to the fact that woodland-dweller early Homo was forced to disperse to the north after the spread of dry savannas in Eastern Africa at about 1.8 million years ago. In this way, the Levant Corridor could have acted as a gate. This gate was shut or hard to cross during the periods of interglacial climate due to the spread of desert conditions in the area, as at present, and opened with expansion of forest precisely at the peaks of glaciation. This is exactly what happened during the climate crisis at 1.8 million years ago. While the savannah and the desert expanded and the forests shrank in sub-Saharan Africa, north of the Sahara, the forests spread along the Levant Corridor, linking up with the Taurus and Zagros mountain forests and the moist relief in the Caucasus. The fortunate hominin populations at the northern extreme of the Rift were able to perpetuate their way of life in these woodland areas further north, spreading naturally into the southern Caucasus and leaving behind the east African savannahs.
REFERENCES
Agustí, J., & Antón, M. (2002). Mammoths, sabertooths, and hominids. New York: Columbia University Press.
Agustí, J., & Lordkipanidze, D. (2011). How «African» was the early human dispersal out of Africa? Quaternary Science Review, 30(11–12), 1338–1342. doi: 10.1016/j.quascirev.2010.04.012
Arribas, A., & Palmqvist, P. (1999). On the ecological connection between sabre-tooths and hominids: Faunal dispersal events in the Lower Pleistocene and a review of the evidence for the first human arrival in Europe. Journal of Archaeological Science, 26, 571–585. doi: 10.1006/jasc.1998.0346
Blumenschine, R. J. (1988). An experimental model of the timing of hominid and carnivore influence on archaeological bone assemblages. Journal of Archaeological Science, 15, 483–502. doi: 10.1016/0305-4403(88)90078-7
Blumenschine, R. J., & Marean, C. W. (1993). A carnivore’s view of archaeological bone assemblages. In J. Hudson (Ed.), From bones to behaviour: Ethnoarchaeological and experimental contributions to the interpretation of faunal remains (pp. 273–300). Carbondale: Southern Illinois University.
Carbonell, E., Mosquera, M., Rodríguez, X. P., Sala, R., & Van der Made, J. (1999). Out of Africa: The dispersal of the earliest technical systems reconsidered. Journal of Anthropological Archeology, 18(2), 119–136. doi: 10.1006/jaar.1998.0331
Dennell, R. (2009). The Palaeolithic settlement of Asia. Cambridge: Cambridge University Press.
Domínguez-Rodrigo, M. (2001). A study of carnivore competition in riparian and open habitats of modern savannas and its implications for hominid behavioral modelling. Journal of Human Evolution, 40, 77–98. doi: 10.1006/jhev.2000.0441
Horowitz, A. (1989). Continuous pollen diagrams for the last 3.5 m.y. from Israel: Vegetation, climate and correlation with the oxygen isotope record. Palaeogeography, Palaeoclimatology, Palaeoecology, 72, 63–78. doi: 10.1016/0031-0182(89)90132-6
Lordkipanidze, D., Jashashvili, T., Vekua, A., De León, M. S. P., Zollikofer, C. P. E., Rightmire, G. P., … Rook, L. (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, 449, 305–310. doi: 10.1038/nature06134
Lordkipanidze, D., Ponce de León, M., Margvelashvili, A., Rak, Y., Rightmire, G. P., Vekua, A., & Zollikofer, C. P. E. (2013). A complete skull of early Homo from Dmanisi, Georgia, and its implications for the evolution and population biology of the genus Homo. Science, 342(6156), 326–331. doi: 10.1126/science.1238484
Marean, C. W. (1989). Sabertooth cats and their relevance for early hominid diet and evolution. Journal of Human Evolution, 18(6), 559–582. doi: 10.1016/0047-2484(89)90018-3
Marean, C. W., & Erhardt, C. L. (1995). Paleoanthropological and paleoecological implications of the taphonomy of a sabertooth’s den. Journal of Human Evolution, 29(6), 515–547. doi: 10.1006/jhev.1995.1074
Martínez-Navarro, B., & Palmqvist, P. (1995). Presence of the African Machairodont Megantereon whitei (Broom, 1937) (Felidae, Carnivora, Mammalia) in the Lower Pleistocene of Venta Micena (Orce, Granada, Spain), with some considerations on the origin, evolution and dispersal of the genus. Journal of Archaeological Science, 22, 569–582. doi: 10.1006/jasc.1994.0054
Shatilova, I., Mchedlishvil, N., Rukhadze, L., & Kvavadze, E. (2011). The history of the flora and vegetation of Georgia (South Caucasus). Tbilisi: Institute of Paleobiology. Georgian National Museum.
Tappen, M., Lordkipanidze, D., Bukshianidze, M., & Ferring, R. (2007). Are you in or out (of Africa?). In T. R. Pickering, K. Schick, & N. Toth (Eds.), Breathing life into fossils: Taphonomic studies in honor of C. K. Brain (pp. 119–135). Bloomington, IN: Stone Age Institute Press.
Tchernov, E. (1992). The Afro-Arabian component in the Levantine mammalian fauna: A short biogeographical review. Israel Journal of Zoology, 38, 155–192.
Vekua, A., Lordkipanidze, D., Rightmire, G.P., Agustí, J., Ferring, R., Maisuradze, G., Mouskhelishvili, A., … Zollikofer, C. (2002). A new skull of early Homo from Dmanisi, Georgia. Science, 297, 85–89. doi: 10.1126/science.1072953




