

Virtual sauropods
A revolution in the study of long-necked dinosaurs

Over the past 20 years, the study of fossils using 3D visualisation techniques, better known as virtual palaeontology, has revolutionised the study of organisms from the past. The study of the popular sauropod dinosaurs is a clear example: the complex mounting systems or expensive replicas once used to study their skeletons are now easily achievable thanks to advances in computing. Thus, questions such as the position of their long necks, their movement capabilities and functional implications, or even how they mated, are easier to address than ever before. Therefore, the insights gained through traditional methods, together with the contributions of new technologies, are opening up new frontiers in the study of these titans.
Keywords: virtual palaeontology, range of neck movement, copulation, giraffes, Jurassic.
Introduction: A revolution in palaeontology
Sauropods were dinosaurs with a distinctive body plan: quadrupeds with long tails and necks that grew to enormous sizes. The largest reached the height of four-storey buildings and had masses of up to 70 tonnes (as much as 250 elephants), making them the biggest animals ever to walk the Earth (Hallett & Wedel, 2016; Sander et al., 2010). Since the first reasonably complete skeletons were discovered in the late 19th century, there have been many interpretations of how these impressive animals fared in their prehistoric environment, in a debate that continues to this day (Taylor, 2010). For decades they were considered to be aquatic due to their large size, but by the end of the 20th century they were interpreted as reptilian giraffes: phytophagous (plant-eating) animals with imposing necks crowned by small heads that fed from the treetops (Bakker, 1971). However, the study of fossils using 3D visualisation, commonly known as virtual palaeontology, has revolutionised not only the study of fossils in general (Sutton et al., 2014), but also the interpretation of sauropod dinosaurs since the early 2000s (Stevens, 2013; Stevens & Parrish, 1999; Taylor et al., 2009; Vidal, Mocho, Aberasturi et al., 2020; Vidal, Mocho, Páramo et al., 2020).
The neck posture debate
Rapid advances in computer technology have made it possible to extract unprecedented information from dinosaur fossils. In the case of sauropod dinosaurs, this technology has enabled the study of features ranging from their bone cavities (e.g., those that housed their central nervous or auditory systems) without the need to physically alter them to something as simple yet revolutionary as working in a virtual environment with dozens of bones simultaneously (something complicated in the physical world given that they weight hundreds of kilos each).
Early on, this technology enabled work to compare the necks of the popular sauropods Apatosaurus and Diplodocus to measure their range of motion (Stevens & Parrish, 1999). This study digitised the articular facets of the cervical vertebrae of these dinosaurs from illustrations of the bones published in monographic descriptions of these taxa. Then they used those digitised articular facets to calculate the amount of movement the necks were capable of. The results showed that neither Apatosaurus nor Diplodocus had necks as flexible as those of modern giraffes, nor would they have been able to raise their necks far above their shoulders (Stevens & Parrish, 1999). Subsequent studies seemed to support this hypothesis, also showing that the vertebral columns of many sauropod dinosaur species were articulated almost straight, with no neck elevation at the base as there is in many theropods (Stevens & Parrish, 2005).
The conclusion was clear: the image of sauropod dinosaurs as reptilian giraffes feeding from the treetops had to be revised. In addition, new fossil finds, such as the small Nigersaurus from 110 million years ago, showed clear adaptations to feeding on vegetation at ground level, such as a downward facing skull, short forelimbs, and a dental battery with dozens of replacement teeth (Sereno et al., 2007). These findings soon influenced the interpretation of the vast majority of sauropods to that of animals with very inflexible horizontal necks, almost incapable of looking behind them or lifting them above their shoulders. This led to depictions such as the Diplodocus specimens in the famous 1999 BBC documentary Walking with dinosaurs.
However, this popular hypothesis of sauropods with less flexible and more or less horizontal necks has not been unanimously accepted by the scientific community. Indeed, it has been shown that modern terrestrial vertebrates, from rabbits to camels and crocodiles, typically hold their necks in a higher position than suggested by virtual palaeontological data (Taylor et al., 2009). Therefore, does this mean that virtual models are too limited to infer the capabilities of vertebrates?
Virtual Spinophorosaurus: a new change of interpretation
Studies of Spinophorosaurus nigerensis, an exceptionally complete and well-preserved early sauropod from the Jurassic period of Niger (from almost 170 million years ago), have brought the two lines of evidence together and shown that, far from being incompatible, they are perfectly complementary. Spinophorosaurus is known from three articulated specimens (Remes et al., 2009): a spectacular, almost complete, articulated skeleton of an 11-metre-long subadult; partial remains of a 13-metre-long adult; and the neck and first back vertebrae of a 3-metre-long juvenile.
Unlike previous work, where two-dimensional images were digitised to create a three-dimensional projection, the bones of these three Spinophorosaurus were all digitised individually using a technique called digital photogrammetry to create a complete virtual Spinophorosaurus skeleton. This technique, based on the same basic principle as a stereoscope, involves taking many slightly overlapping photographs that capture the entire surface of each fossil bone in the skeleton (Mallison & Wings, 2014). Computer processing produces a very large number of homologous points between all the photographs, which are used to construct a virtual surface that reproduces the fossil with a margin of error of less than a millimetre – provided the scanning is performed correctly. By repeating this process for each bone, a collection of virtual bones ready for articulation is obtained.
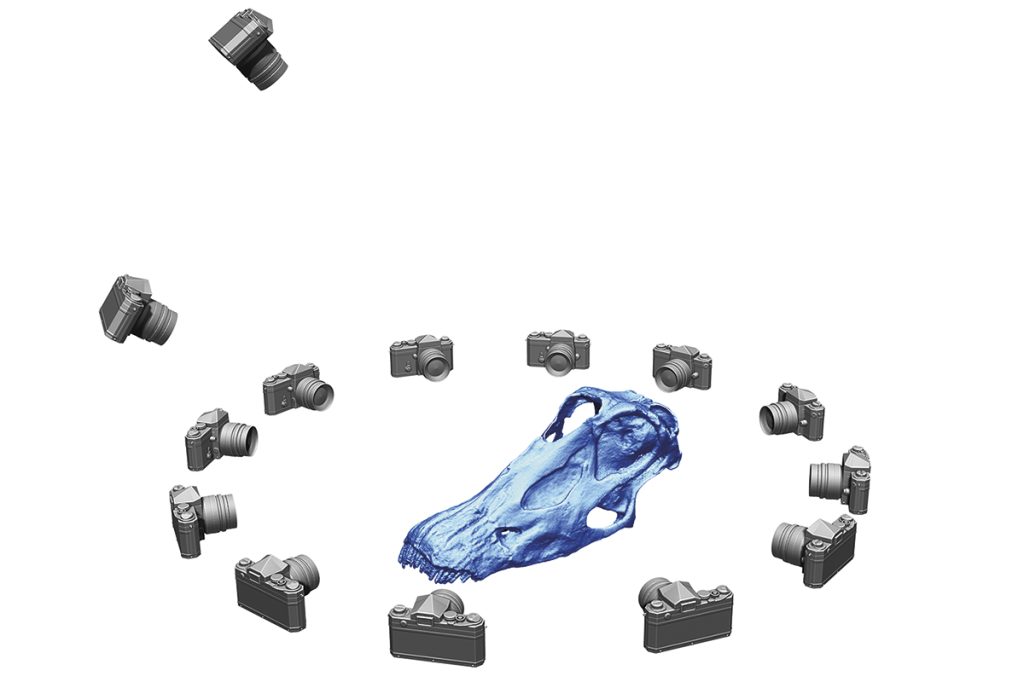
Schematic of the process of digitising a Diplodocus skull using digital photogrammetry, in which a large number of slightly overlapping photographs that cover the entire surface of the fossil are taken. These images are then processed in a computer to obtain a large number of homologous points between all the photographs, which are used to generate a virtual surface. / Daniel Vidal
The first step in getting an idea on how a vertebrate skeleton is arranged is to articulate the bones in what is called the osteological neutral position, the position that results from articulating the bones at the maximum contact allowed by their facet joints. Although it is called neutral, it does not necessarily mean that it was the most common posture adopted by the animal in life (Dzemski & Christian, 2007). However, it is a relatively objective starting point that allows the skeletal arrangement of different vertebrate species to be compared using the same criterion: maximum joint contact. In an attempt to avoid preconceived notions of what the osteologically neutral posture might be, the spine is articulated with only one pair of vertebrae visible at a time. With one vertebra completely static, the next vertebra is articulated with all its facet joints in contact. Once these first two vertebrae have been articulated, the software allows to turn the first vertebra invisible and import the following vertebra. Thus, only the second and third bones can then be seen and the position of the third bone can be adjusted so that it fits perfectly with the second one. This process removes any bias that preconceived notions might introduce into the neutral posture of the spine. Nonetheless, these preconceptions still strongly influence how sauropod skeletons are articulated in many museums, where the disarticulation of the spine is often forced in several places to achieve, for example, a curved back posture.
Surprisingly, the virtual skeleton resulting from applying this protocol was radically different from all the previous reconstructions of this dinosaur species, which traditionally showed Spinophorosaurus with a horizontal vertebral column. The new reconstruction showed an upward sloping neck and back with a horizontal tail (Vidal, Mocho, Aberasturi et al., 2020; Vidal, Mocho, Páramo et al., 2020). Despite its elevation, the neck was virtually aligned with the dorsal vertebrae, with no significant change in the angle of elevation at the base of the neck, as had been shown in previous studies (Stevens & Parrish, 2005). But how could Spinophorosaurus have had its neck tilted upwards? There had to be a point where the vertebral column rose, where the sauropod «keystone» was found. We found that keystone in the sacral vertebrae, the vertebrae that had fused with the hip bones.
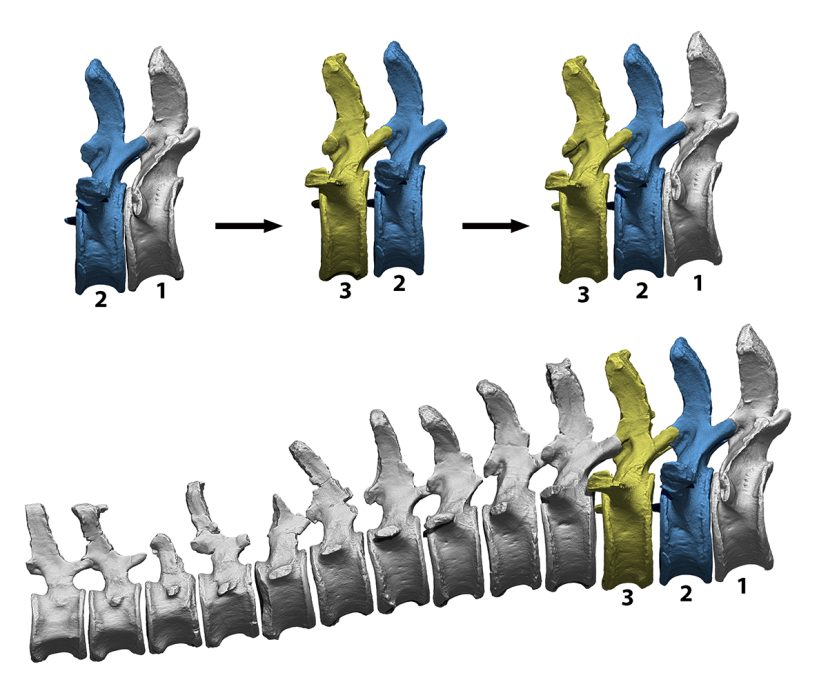
The process of articulating a sequence of a virtual caudal vertebrae of the sauropod Camarasaurus in a neutral posture. This technique is used to prevent preconceived ideas from having a significant effect on the eventual outcome of the curvature of the spine. First, the first pair of vertebrae is articulated to maximise the overlap of their articular surfaces. Then the first caudal vertebra is hidden and the third one is added, with only one pair of vertebrae visible at a time. The resulting curvature is the result of maximum articular contact, in case there were potential biases in interpretation. / Daniel Vidal
This was surprising because most palaeontologists expected the keystone to be where the neck met the back or the base of the neck (Stevens, 2013; Taylor et al., 2009). However, as the palaeontologist Henry F. Osborn (1899) noted, just as in birds, «the sacro-iliac junction is the center of power and motion» and is the most reinforced area of the spine. Therefore, this area could withstand changes without compromising the animal’s ability to walk. The sacrum of Spinophorosaurus is wedge-shaped: it is longer in the lower half and shorter in the upper half. As a result, when the tail is articulated, the torso and neck were raised, allowing Spinophorosaurus to raise its snout five metres above the ground from this neutral position, more than twice the height of its shoulder.
Were sauropods like giraffes?
To find out whether these techniques are really capable of providing accurate data on sauropod palaeobiology, the most immediate approach is to assess whether they allow us to replicate the biological capabilities of modern vertebrates. Giraffes are the only vertebrates today that can be used as a model for the necks of sauropods: they are large quadrupeds that weigh over a tonne and have a long neck as a result of the elongation of their cervical vertebrae and the «cervicalisation» of their first thoracic vertebra. Notably, by applying the same techniques described for sauropods to modern giraffe skeletons, the skeletal reconstructions obtained correspond perfectly to the postures normally adopted by giraffes when walking (Vidal, Mocho, Páramo et al., 2020).
In the wild, giraffes usually feed with their necks above their shoulders, but occasionally males feed on the tallest shoots they can reach with their necks fully erect, and females feed by grazing at ground level. All this range of giraffe neck positions can be achieved with their virtual skeletons without the need to disarticulate any vertebrae. Moreover, even the maximum ventroflexion position of the neck of virtual giraffes does not allow them to reach the ground without bending their elbows or spreading their shoulders, as is the case with live giraffes. The data shows that the entire repertoire of giraffe neck positions can be replicated virtually using the same parameters as for sauropods.
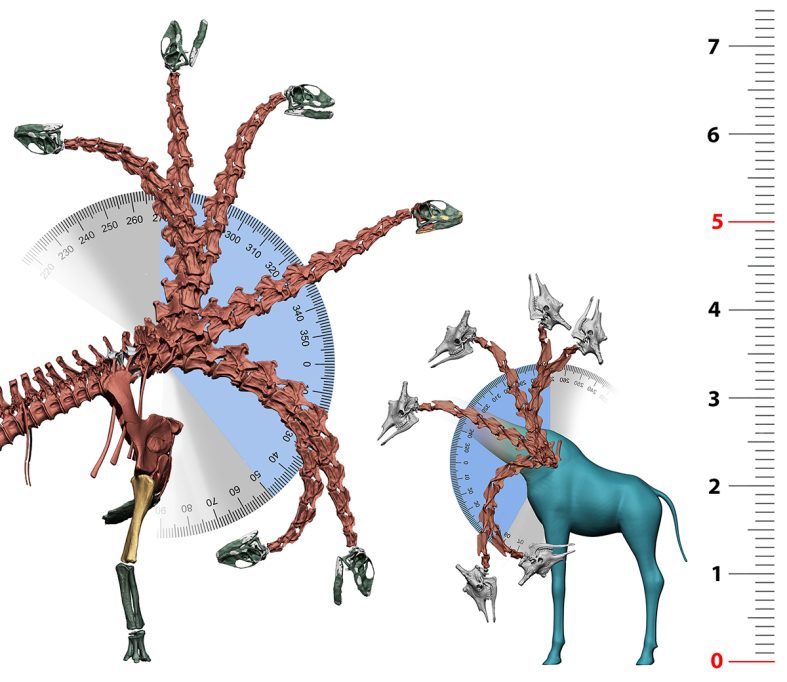
The range of motion of the neck of the sauropod dinosaur Spinophorosaurus nigerensis is remarkably similar to that of the modern giraffe. The enormous size of Spinophorosaurus allowed it to reach heights of more than 7 metres simply by bending its neck, making it one of the earliest sauropod dinosaurs known to be able to position its head at this height. / Daniel Vidal
So, what does this mean for sauropods? The answer is more complex than saying that if an extinct organism could physically adopt a posture, then it was a habitual posture and part of its normal behaviour. We humans can walk on our toes when ballet dancing, and elephants can stand on their forelimbs, but this does not mean that ballet dancing or forelimb standing are common behaviours in either species (Mallison, 2010; Sander et al., 2010). However, the ability to do so does exist. Virtual palaeontology techniques can reveal the physical capabilities of extinct vertebrates and which postures or behaviours were physically impossible for them and should therefore be discarded when interpreting the ecomorphology of these organisms (Mallison, 2010). In the case of Spinophorosaurus nigerensis, its skeleton allowed positions that the animal could have adopted to feed the same way as extant giraffes: from treetops almost 8 metres high to vegetation at ground level. It is one of the earliest sauropods for which we have evidence of these abilities. Other sauropods, such as the diplodocoids Apatosaurus, Diplodocus, or Amargasaurus, did not have skeletons that gave them the same movement abilities and were more restricted in the vertical positioning of the neck, suggesting that they probably occupied different ecological niches (Christian et al., 2005; Stevens & Parrish, 1999, 2005). Therefore, virtual models allow us to reproduce the functional capabilities of modern vertebrates and, consequently, to infer those of extinct vertebrates. However, it is impossible to know whether all these capabilities were part of their behaviour or, if they were, whether they were frequent or occasional. In any case, these techniques can be used to refute behaviours when they are incompatible with the capabilities of an organism.
Mating in sauropods
The correct interpretation of how non-avian dinosaurs reproduced is a complicated mystery. We know for a fact that dinosaurs, even the largest sauropods, could mate because they all had offspring and descendants, but their gigantic size makes us wonder what exactly this behaviour was like. To get an idea of how sauropod dinosaurs mated, we first need to look at how their closest living relatives, crocodiles and birds, copulate. Copulation in crocodilians is remarkably similar in all extant species and is the same as in many reptiles: the male stands next to the female and places his hind leg on the female’s hip, a position called leg over back. This position makes it easier for the male’s «penis» to enter the female’s cloaca, as the caudal appendage of reptiles often makes copulation difficult (Isles, 2009). Evolution has greatly reduced the caudal appendage size of birds, which in most modern species consists of a few mobile vertebrae that do not interfere with copulation. This is why birds do not adopt the leg-over-back posture of crocodilians. Most basal birds have a «penis» (similar in function to that of mammals, although of a different evolutionary origin), but passerines and galliformes tend to reduce or lose the caudal appendage entirely (Isles, 2009).
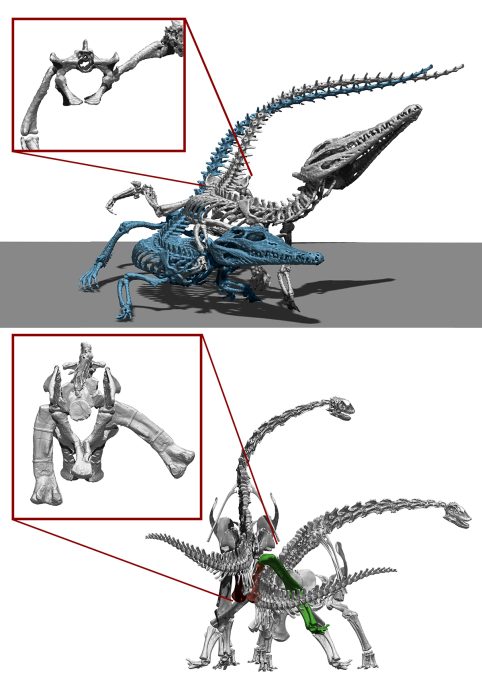
Modern crocodilians have the ability to lift their hind leg in abduction well above the horizontal plane, allowing them to copulate in a leg-over-back position. However, sauropod dinosaurs could barely perform such abductions because the proximal epiphysis of the femur would have collided with the walls of the acetabulum. This means that sauropods were not capable of copulation in the same way as most modern reptiles. / Daniel Vidal
The fact that the body plan of crocodilians is more similar to that of sauropods (they are quadrupeds and have a well-developed tail appendage) suggests that the latter may also have adopted the leg over back position as a strategy to facilitate copulation (Hallet & Wedel 2016; Isles, 2009). However, the skeleton of sauropods would simply not allow this position. Unlike crocodilians, which have a femur with a rounded head that allows a lot of mobility in the hip socket, sauropods have femora with an inverted L-shaped head. This means that any attempt at abduction is soon limited by collision with the walls of the acetabulum, greatly restricting movement in this plane. The limited ability of sauropods to elevate their hind limbs means that there would not have been enough space between the hip and the hind limb to accommodate the body of the female, as is the case in crocodilians, unless the female was much smaller than the male. There are few studies of sexual dimorphism in sauropods, but the little evidence available suggests that there was no marked size dimorphism between the sexes (Ikejiri, 2004). Therefore, it is safe to say that sauropods could not have adopted the same copulatory position adopted by crocodiles and by the vast majority of modern reptiles, and that their copulatory strategies must have been different. Other possibilities are that sauropods could have adopted a mating pattern similar to that of most quadrupedal mammals, with the male standing on his hind limbs and supporting himself with his forelimbs on the female, who could be standing or lying down. There is nothing in the osteological range of motion of sauropods that would have prevented them from adopting these positions. However, there is also no osteological impediment to them having copulated back-to-back, as many arthropods and some canids do. Ultimately, we cannot know how sauropods mated because we cannot rule out all these possibilities. What we can say with absolute certainty is that they did not do it in the same way as today’s crocodiles.
Conclusion: An additional and complementary tool
Having seen these examples, it is clear that virtual technologies are not a new field that has come to replace traditional palaeontology and comparative anatomy because the latter have become obsolete. Rather, these new technologies are an additional and complementary tool that can scientifically evaluate certain hypotheses that were previously exceedingly difficult or even impossible to test. We have seen how virtual palaeontology shows that the traditional hypothesis of sauropods as giraffe analogues requires much more nuance. Some sauropods had giraffe-like feeding capabilities, while others had more limited neck mobility, which conditioned them to feed on low-growing or ground-level vegetation. This suggests a more complex scenario than previously thought, in which the question is not whether all sauropods could feed like giraffes, but rather, which ones could or could not do this.
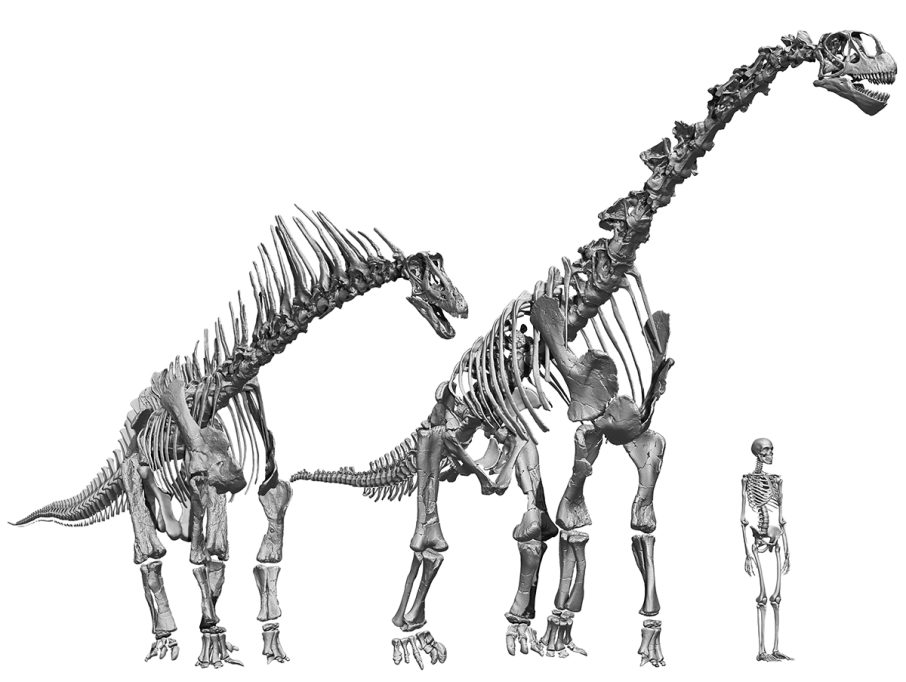
Different sauropods had osteologically neutral postures and neck movements, which probably resulted in different ecomorphologies. Many diplodocoids, such as Amargasaurus, had necks with fairly horizontal neutral postures which has been interpreted as them having been foragers of medium, low, and ground vegetation. In contrast, macronarian sauropods such as Camarasaurus had necks with a much higher neutral posture and are therefore interpreted as having been foragers of taller vegetation. / Daniel Vidal
Similarly, until recently, how sauropods mated has been a matter of speculation. However, thanks to virtual palaeontology, it has been possible to disprove the idea that sauropods mated in the same way as modern crocodiles. Virtual palaeontology has always required observations of living animals or the availability of exceptionally complete fossil specimens to be able to evaluate these hypotheses convincingly. It is both a revolution in the way we study dinosaur fossils and a reminder of the importance of combining multiple lines of evidence as the best way to advance scientific knowledge.
Bakker, R. T. (1971). Ecology of the brontosaurs. Nature, 229, 172–174. https://doi.org/10.1038/229172a0
Dzemski, G., & Christian, A. (2007). Flexibility along the neck of the ostrich (Struthio camelus) and consequences for the reconstruction of dinosaurs with extreme neck length. Journal of Morphology, 268, 701–714. https://doi.org/10.1002/jmor.10542
Hallett, M., & Wedel, M. J. (2016). The sauropod dinosaurs. Johns Hopkins University Press.
Ikejiri, T. (2004). Anatomy of Camarasaurus lentus (Dinosauria: Sauropoda) from the Morrison formation (Late Jurassic), Thermopolis, Central Wyoming, with determination and interpretation of ontogenetic, sexual dimorphic and individual variation in the genus. Fort Hays State University.
Isles, T. E. (2009). The socio-sexual behaviour of extant archosaurs: Implications for understanding dinosaur behaviour. Historical Biology, 21(3–4), 139–214. https://doi.org/10.1080/08912960903450505
Mallison, H. (2010). CAD assessment of the posture and range of motion of Kentrosaurus aethiopicus Hennig 1915. Swiss Journal of Geosciences, 103(2), 211–233. https://doi.org/10.1007/s00015-010-0024-2
Mallison, H., & Wings, O. (2014). Photogrammetry in paleontology–A practical guide. Journal of Paleontological Techniques, 12, 1–31.
Osborn, H. F. (1899). A skeleton of Diplodocus, recently mounted in the American Museum. Science, 10(259), 8070–874. https://doi.org/10.1126/science.10.259.870
Remes, K., Ortega, F., Fierro, I., Joger, U., Kosma, R., Ferrer, J. M. M., Ide, O. A., & Maga, A. (2009). A new basal sauropod dinosaur from the middle Jurassic of Niger and the early evolution of Sauropoda. PLOS ONE, 4(9), e6924. https://doi.org/10.1371/journal.pone.0006924
Sander, P. M., Christian, A., Clauss, M., Fechner, R., Gee, C. T., Griebler, E.-M., Gunga, H.-C., Hummel, J., Mallison, H., Perry, S. F., Preuschoft, H., Rauhut, O. W. M., Remes, K., Tutken, T., Wings, O., & Witzel, U. (2010). Biology of the sauropod dinosaurs: The evolution of gigantism. Biological Reviews, 86, 117–155. https://doi.org/10.1111/j.1469-185X.2010.00137.x
Sereno, P. C., Wilson, J. A., Witmer, L. M., Whitlock, J. A., Maga, A., Ide, O., & Rowe, T. A. (2007). Structural extremes in a cretaceous dinosaur. PLOS ONE, 2(11), 1–9. https://doi.org/10.1371/journal.pone.0001230
Stevens, K. A. (2013). The articulation of sauropod necks: Methodology and mythology. PLOS ONE, 8(10), 1–27. https://doi.org/10.1371/journal.pone.0078572
Stevens, K. A., & Parrish, M. J. (1999). Neck posture and feeding habits of two Jurassic sauropod dinosaurs. Science, 284, 798–800. https://doi.org/
10.1126/science.284.5415.798
Stevens, K. A., & Parrish, M. J. (2005). Neck posture, dentition, and feeding strategies in Jurassic sauropod dinosaurs. In V. Tidwell & K. Carpenter (Eds.), Thunder-lizards: The sauropodomorph dinosaurs (pp. 212–232). Indiana University Press.
Sutton, M. D., Rahman, I. A., & Garwood, R. J. (2014). Techniques for virtual palaeontology. Wiley. https://doi.org/10.1002/9781118591192
Taylor, M. P. (2010). Sauropod dinosaur research: A historical review. Geological Society London Special Publications, 343(1), 361–386. https://doi.org/10.1144/SP343.22
Taylor, M. P., Wedel, M. J., & Naish, D. (2009). Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica, 54(2), 213–220. https://doi.org/10.4202/app.2009.0007
Vidal, D., Mocho, P., Aberasturi, A., Sanz, J. L., & Ortega, F. (2020). High browsing skeletal adaptations in Spinophorosaurus reveal an evolutionary innovation in sauropod dinosaurs. Scientific Reports, 10(1), 6638. https://doi.org/10.1038/s41598-020-63439-0
Vidal, D., Mocho, P., Páramo, A., Sanz, J. L., & Ortega, F. (2020). Ontogenetic similarities between giraffe and sauropod neck osteological mobility. PLOS ONE, 15(1). https://doi.org/10.1371/journal.pone.0227537




