History and reality of the genus ‘Homo’
What is it and why do we think so?
Paleoanthropologists who worry about how nature is organized into species, and about what we should call them, are very often accused by their peers of «just arguing about names». This implies that basic taxonomy is a boring clerical operation that should be dispensed with as quickly as possible or even ignored, so that we can get to the really interesting questions about human evolution. Yet the reality is that we shall never understand the events of the intricate human evolutionary play if we cannot accurately identify the actors who participated in that drama. This article looks briefly at how our current supremely woolly concept of the genus Homo has come about, as background for urging a more rational approach to defining it.
Keywords: taxonomy, genera, genus Homo, palaeoanthropology.
The genus ‘Homo’: Framing the question
The first issue that arises in relation to defining the genus Homo is the nature of the unit itself. Nowadays most would agree that in principle genera should be not only monophyletic but holophyletic. That is to say, a genus should contain all of the species descended from a particular common ancestor – and only those species. But there is no rule, even an empirical one, to say how many species should exist within a genus, or how remote that common ancestor can acceptably be. What is more, some well-established living genera are rich in species, while others are much more species-poor, and some contain only one. Partly this variety occurs because some genera appear inherently more disposed than others to diversification; but in part it is also due to a taxonomic flexibility that recognizes there is no «natural» boundary for a genus – as there is, at least in theory, for species.
«A genus should contain all of the species descended from a particular common ancestor, and only those species»
Fortunately, in the living world genera tend to be fairly readily recognizable as Gestalt units.1 This is because morphological gaps between related genera are typically much greater than those separating species within the same genus, since as the taxonomic net widens, differences naturally accumulate, even if at irregular rates. In the fossil record taxonomic judgments are inherently trickier than they are among living forms, not only because the information available is inevitably more limited, but also because the evidence we do have tends to be rather fragmentary, and scattered over time and place.
Our current understanding in every branch of science is deeply conditioned by the particular history of each field. And in the case of palaeoanthropology it is especially true that today’s received wisdom is – to an alarming extent – the product of the entirely arbitrary sequence in which the components of the known fossil record were discovered.
Our current paleoanthropological beliefs have also been deeply affected over the past couple of centuries by huge fluctuations in our ideas about what genera are and should be. Because of this reality, the consideration of the genus Homo that follows is largely a historical one. For almost certainly, if the entire hominid fossil record were to be discovered tomorrow, rather than in dribs and drabs over a century and a half, we would view it very differently than we do today.
Early Notions of the Genus ‘Homo’
Linnaeus’ original concept of the genus Homo, articulated in 1758 (Linnaeus, 1758) was most closely comparable to today’s superfamily Hominoidea, for it embraced not only Homo sapiens but Homo troglodytes, a species that included both chimpanzees and orangutans. The notion that Homo sapiens might have shared its genus with now-extinct relatives did not surface until 1864, when William King classified as Homo neanderthalensis a clearly ancient partial fossil skeleton that had been found in Germany’s Neander Valley (Thal in German) (King, 1864). For three decades, though, that was about it as far as naming new extinct forms of Homo was concerned, principally because discussion of humanity’s biological origins had to be carried on in the absence of any fossils apart from a few that resembled the Neanderthaler. Not that everyone found this inhibiting.
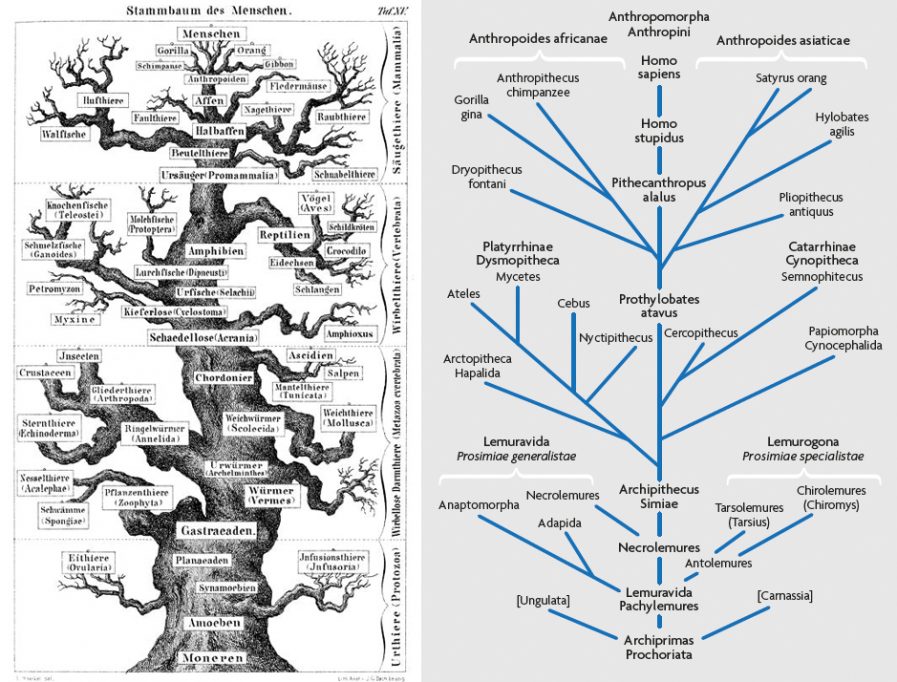
Figure 1 shows Ernst Haeckel’s notion of the tree of life, published in 1868 when there was, apart from the Neanderthaler itself, virtually no human fossil record to speak of. Haeckel’s drawing gives us the classic image of a tree, with a tapering if gnarly trunk, and Homo sapiens at its apex. In a later and more detailed phylogeny of the primates (Figure 2), Haeckel went even further, although he could only imagine at that point what the fossil history of humankind might reveal. But he knew that there had to have been earlier links connecting human beings to the rest of nature, so he inserted into his phylogeny the two hypothetical species Homo stupidus (not the Neanderthaler, in case you’re wondering) and Pithecanthropus alalus, in a linear fashion of direct ancestry and descent (Haeckel, 1868).
Thus it was that in 1891, when Eugene Dubois discovered, at Trinil in Java, the type calotte of the fossil hominid species he eventually called Pithecanthropus erectus, it made perfect sense for him to regard this specimen, which was smallish-brained yet was clearly in some sense human, as a lineal ancestor of Homo sapiens. This amazing find seemed to fit perfectly into Haeckel’s construct; so perfectly, indeed, that Dubois appropriated for his fossil Haeckel’s genus name Pithecanthropus (“ape-man”). This intermediate placement is reflected in Dubois’s phylogeny diagram of 1896 (Figure 3; see Theunissen, 1988). And, thanks to the efforts not only of Dubois but of many other influential scholars such as the German anatomist Gustav Schwalbe, by the turn of the twentieth century it was widely agreed that Dubois’ Pithecanthropus erectus was the ancestral species that had given rise to Homo sapiens.
«The notion that ‘Homo sapiens’ might have shared its genus with now-extinct relatives did not surface until 1864»
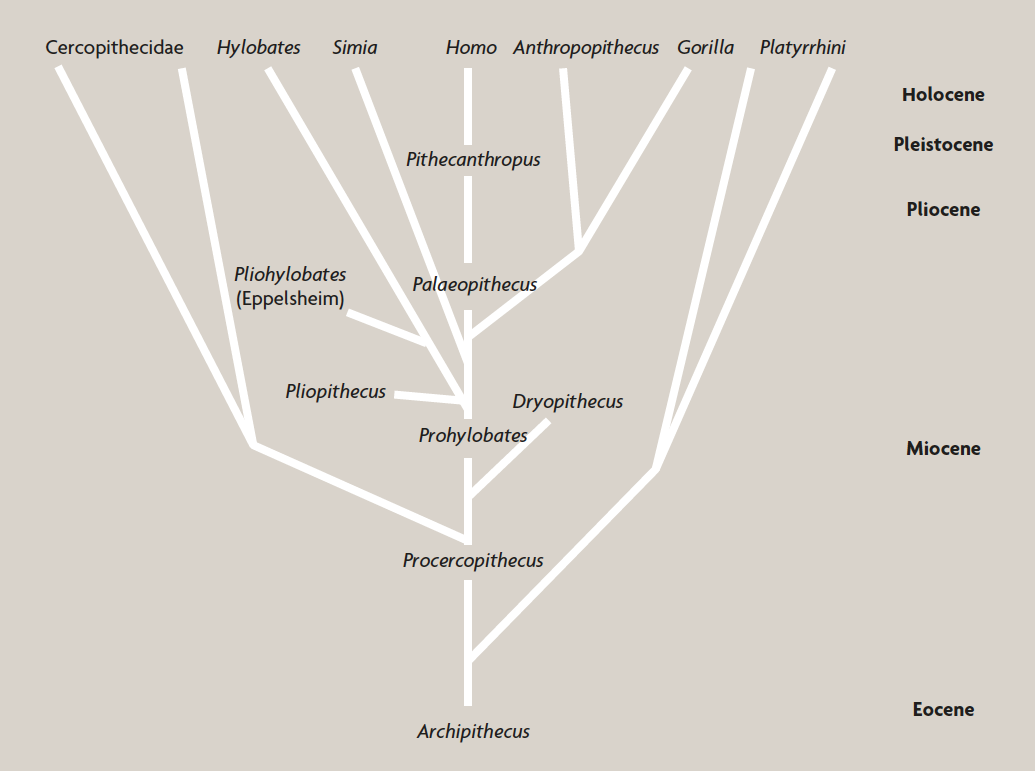
But surprisingly soon, and perhaps inevitably, interpretations began to change. By the beginning of the second decade of the twentieth century, Dubois’ Pithecanthropus had once more become distinctly peripheralized, as seen in Figure 4, a human phylogeny diagram published by the British anatomist Sir Arthur Keith (1915). In this vision, virtually all the human fossils or fossil clusters then known were seen as parts, or endpoints, of terminal lineages. Significantly, most of those writing about the human fossil record during that period were human anatomists like Keith: scientists deeply versed in the minutiae of variation within Homo sapiens, but with little interest in the debates over evolutionary mechanism that raged among geneticists, embryologists, naturalists, and others during the decades following the rediscovery of the principles of Mendelian genetics in 1900. Paleoanthropologists of this period seem to have been content in general to accept Darwin’s notion of evolution as «descent with modification», and to leave it to others to worry about the underlying processes. Their business was simply to examine the results. However, unlike the obligation that so clearly faced comparative anatomists, this did not necessarily mean having to sort out the diversity of the living world. As a result, as far as paleoanthropologists were concerned, names, even formal zoological names, were little more than convenient identifiers for individual fossils, rather than a means of delineating units in nature. Remarkably, this applied as much to genera as to species: to most paleoanthropologists in this period, ours was a genus that contained only Homo sapiens and extinct hominids of similarly large brain size: something that usually meant including the Neanderthals in the genus Homo, but not much more.
By the time the Second World War came around, then, any discernible pattern in the steadily enlarging human fossil record had become obscured by a thicket of generic and informal names. And regardless of the nomenclature preferred, the typical hominid family tree of the interwar period continued, in good Keithian fashion, to represent hominid evolution as a profusion of terminal branches.
The evolutionary synthesis
The movement known as the evolutionary synthesis was born during the 1930s and 1940s out of a theoretically uneasy (but reductionistically appealing) fusion of mathematical population genetics with speciation theory. Ultimately, it reduced the complexities of the evolutionary process to a simple matter of gradual shifts in gene frequencies within evolving lineages of organisms, under the guiding influence of natural selection. Especially in its later «hardened» versions, the synthesis taught that each species gradually transformed into its successor over vast periods of time. The emergence of genera and higher taxa was seen simply as a passive consequence of repeated transitions of this kind.
«For most, ours was a genus that contained only ‘Homo sapiens’ and extinct hominids of similarly large brain size»
One of the most influential proselytizers for the synthesis was the ornithologist Ernst Mayr, who was more responsible than anyone else for finally bringing evolutionary theory into paleoanthropology. In 1950 Mayr published a paper that literally rewrote the rules of how paleoanthropologists went about their business. To understand quite how huge the revolution was that Mayr brought about, it helps to remember that the relatively few hominid fossils then known were denoted by a profusion of genus and species names. In one fell swoop, Mayr reduced some fifteen genus names to only one. What is more, he reduced literally dozens of species names to just three, yielding this classification of the hominid family:
Family Hominidae
Genus Homo
Species Homo transvaalensis [australopitecs]
Species Homo erectus
Species Homo sapiens [including Neanderthals]
Finally, the genus Homo was back at the center of palaeoanthropology. Indeed, in Mayr’s vision it embraced the entire family Hominidae. What’s more, Mayr placed his three species of the genus Homo within one single evolving lineage, thus returning to the pure linearity of Haeckel’s vision. In proposing this breathtaking oversimplification, Mayr was hugely uncomplimentary about the supposedly «unbiological» stance of the prewar palaeoanthropologists, whom he rightly reproached for neglecting to think in terms of variability within biological populations. Remarkably enough, at least in the English-speaking world paleoanthropologists accepted Mayr’s blunt criticism with amazing meekness. Indeed, they stampeded to join the unilinealist bandwagon, possibly because they had been subliminally aware all along of the theoretical vacuum that had existed at the center of their work. So profound were the effects of Mayr’s attack that, within a decade, hardly anyone was daring to use species or genus names at all.
Mayr’s concern with intra-species variation was, of course, a valid one. And it fit comfortably with the human anatomical tradition of documenting variation within Homo sapiens. But the new focus also meant that nobody was concerned at all with any morphological synapomorphies3 that might have held the newly reborn genus Homo together. So instead, we typically got diagrams like the one in Figure 5 from a classic paper by the American paleoanthropologist Clark Howell (1957).
«The evolutionary synthesis was born of the fusion of mathematical population genetics with speciation theory»
In Howell’s diagram the players in the human evolutionary game are all individual fossils, mostly contained within a single formless phylogenetic blob that gives no impression at all of the nature of diversity among the hominids, or of what might hold the genus Homo together. For years afterwards, paleoanthropologists continued routinely to publish minimalist phylogenies of this kind, with a broad main hominid trunk giving rise at most to one single aberrant and short-lived side lineage: the robust australopiths, which even Mayr (1953) had been forced to sideline after a broadside from John Robinson (1951). In such linear constructs, species – and by extension genera – simply graded insensibly from one into the next over time. And the consequence of this was that it became impossible, pretty much by definition, to recognize species and genera on the basis of derived morphological characters – even had anyone wished to do so. Paleoanthropologists had forgotten the fact – if they had ever really grasped it – that genera, as aggregations of species, are higher taxa, and that the genus Homo consequently needs to be defined, like any other clade, on the basis of synapomorphies.
After the synthesis
Given this background, it is hardly surprising that the 1960s was also the period during which the ultra-minimalist «single-species» notion of human evolution gained wide currency. This notion had weirdly-assorted progenitors. One of them was Franz Weidenreich, the brilliant anatomist and monographer of Peking Man (Weidenreich, 1939). Like most of his contemporaries, Weidenreich was very little burdened by any theory of how evolution actually worked, although he seems to have leaned toward orthogenesis, the notion that evolution is somehow goal-oriented. This was anathema to the other progenitors of the single species hypothesis, Theodosius Dobzhansky (1944) and Ernst Mayr (1950); but all three members of the trio were comfortable with the notion that the acquisition of culture had made the human ecological niche so broad that, even in principle, not more than one human species could ever have existed at a given point in time. And if this were truly the case, it meant that, irrespective of how much they differed from each other anatomically, all hominid fossils of the same time period had to have belonged to the same species.
«The impressive fossil record we have today really is yielding a signal of species diversity in hominid history»
Although at mid-century this point was already hard to argue from fossil morphologies, such was Mayr’s impact on palaeoanthropology that it was not until the middle 1970s, when it was shown that two different kinds of hominid had lived on the very same landscape in northern Kenya just under two million years ago, that paleoanthropologists were once again forced to start thinking in terms of systematic diversity among early hominids. Since that point, the rapidly accumulating human fossil record has made it plain that the history of the human family has not simply involved the gradual perfecting of adaptation within a single major human lineage. Instead, the history of the hominid family has been one of constant evolutionary experimentation, with new species regularly appearing, competing on the ecological stage, and as likely as not becoming extinct. And far from being the tip of a fine tall tree, as envisaged by Haeckel and Mayr, Homo sapiens is merely one terminal twig on a luxuriantly branching bush, the complexities of which are becoming more evident all the time.
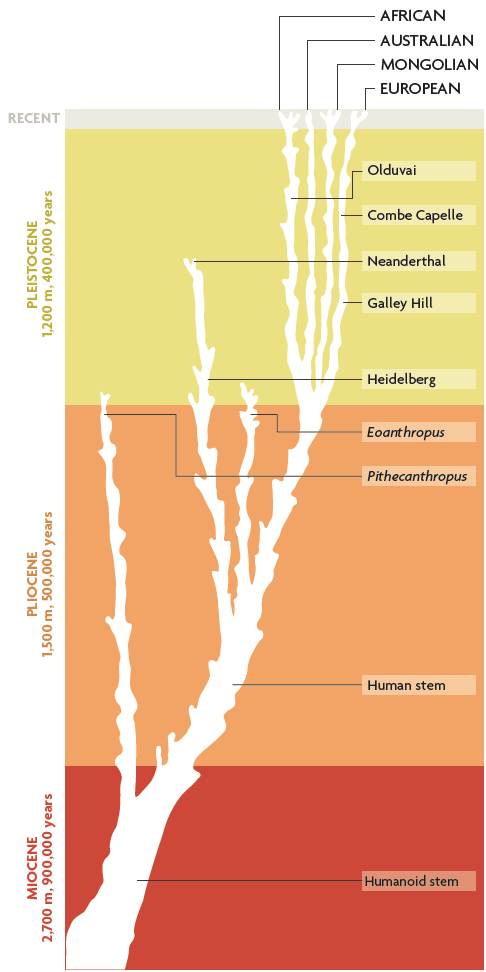
In 1993, I published the very tentative phylogeny of the hominid family (Tattersall, 1993) with eleven hominid species; it was considered fairly extravagant at the time (Figure 6). By the point at which this diagram was redrawn in 1998, the count was up to sixteen. Interestingly, many of the additional species had appeared at the very base of the tree, demonstrating that the pattern of diversity and evolutionary experimentation seen in the original had been present among hominids from the very start. In 2002, the picture had become more complex yet, with twenty species that had somehow to be fitted in; and the most recent version (Figure 7) has 27 species. The details of these very tentative phylogenies, just like the exact number of species at issue here, remain as debatable as they always have been. But what is abundantly clear is that we are not looking at an explosion simply of names on paper. The impressive fossil record we have today really is yielding a signal of species diversity in hominid history.
The genus ‘Homo’ today
This signal of diversity brings us back to the central questions addressed by this article. How is hominid diversity properly organized? How much of it is legitimately contained within our own genus Homo? And how do we recognize the limits of this genus? As already noted, a genus is a grouping of species related by ancestry. But beyond the general requirement of monophyly there is no general criterion, either theoretical or practical, that dictates how inclusive a genus should be. So where do we draw the line?
Despite the best efforts of researchers such as Bernard Wood and Mark Collard (e.g., Collard & Wood, 2015; Wood & Collard, 1999), who have argued strenuously for limiting the genus Homo to hominids both adaptively similar to the type species, Homo sapiens, and more closely related to it than to the type of any other genus, the recent trend has been toward inclusivity. Indeed, the ecumenical genus Homo has now been extended back as far as 2.8 million years (Villmoare et al., 2015). But even assuming that monophyly applies here, to maintain any value as a repository of phylogenetic and/or structural information, the resulting large and inclusive genus Homo would require an elaborate apparatus of subgeneric classification. Which would, of course, defeat any reasonable purpose there might have been for lumping all the hominids concerned together. It thus seems to me to be entirely more practical – as well as more defensible theoretically – to start with Homo sapiens, and to inquire which other hominid species might usefully be classified in a genus that is perforce defined by our own species.
«The 1960s was also the period during which the ultra-minimalist “single-species” notion of human evolution gained wide currency»
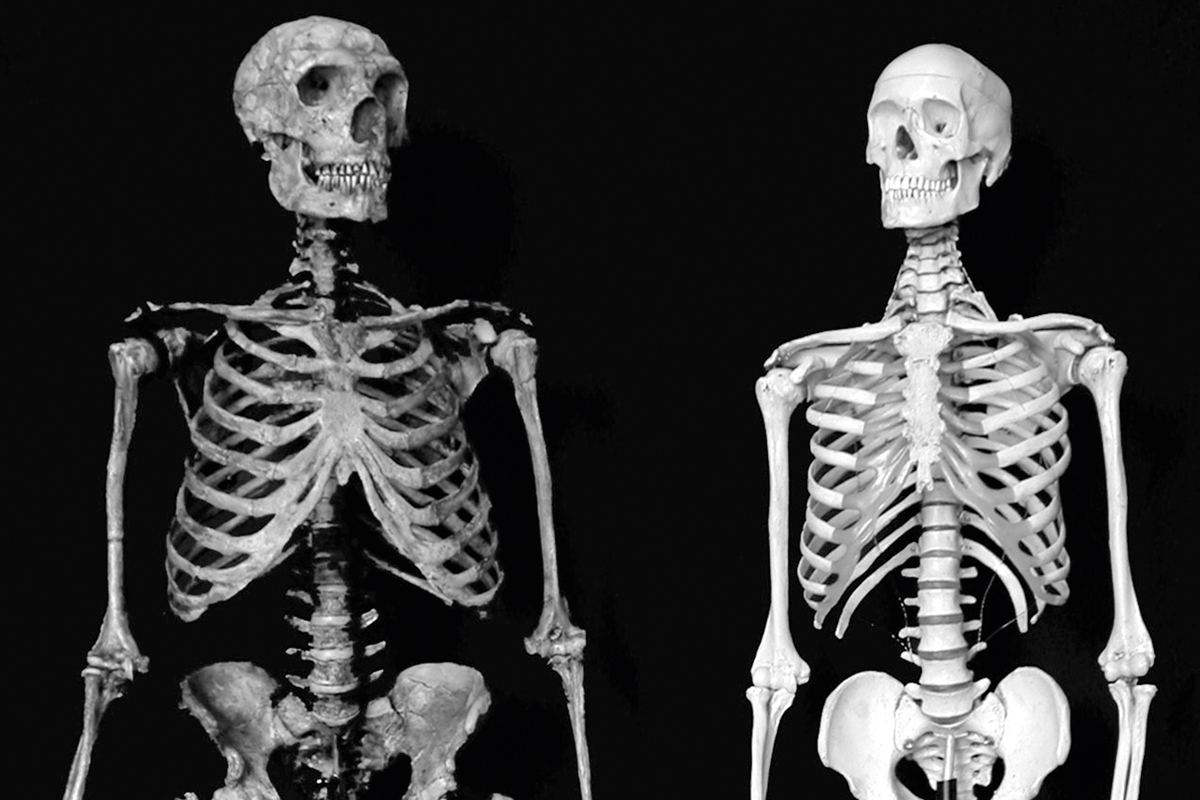
There is, inevitably, a good deal of argument over the number of species that should be recognized among those fossil hominids most closely related to Homo sapiens. For example, it is clear that, within the grouping that used to be loosely known as «archaic Homo sapiens», a variety of different hominids is represented – though few today would see any problem with including all of these forms within the genus Homo. The same is true for the distinctive Neanderthals, currently almost universally recognized as Homo neanderthalensis. These large-brained European hominids are quite closely related to Homo sapiens, even though it is now clear that their clade separated from the lineage leading to Homo sapiens at least a half a million years ago (Arsuaga et al., 2014). It is also worth mentioning in this context that Homo sapiens appears to be quite highly derived in its osteological structure: something that is apparent from the comparison in Figure 8 of a modern human skeleton with a postcranially more primitive composite Neanderthal. Again, though, nobody is presently arguing that any of these differences is a necessary impediment to the inclusion of both in the genus Homo.
Moving outwards yet further, there is active debate over what Homo erectus (the modern incarnation of Dubois’s Pithecanthropus erectus) ought to consist of. One side argues that this species is an Old-World-wide taxon that embraces a vast variety of hominid fossils from Asia and Africa covering almost two million years. Others, including me, would argue that Homo erectus should be regarded as an endemic and terminal Asian form, in which case earlier African hominids often allocated to Homo erectus – but morphologically very dissimilar to the Javan type material – should be placed in their own species, Homo ergaster (Tattersall, 2015a). Once again, though, there seems to be no compelling reason in theory why all members of the erectus/ergaster group should not be viewed as members of a phylogenetically coherent genus Homo. For, although some of them possessed brains of only modest size at best, all seem to have acquired body skeletons of reasonably modern form. In fact, if future analyses and finds confirm that the hominids just mentioned are united by a common postcranial conformation, then we may find it best to recognize the genus Homo not on cranial apomorphies, but on postcranial ones. This may be a bit uncomfortable for modern human beings, who most value themselves for what goes on inside their heads. But it is just possible that, in the end, it will be the only way in which we can recognize a genus Homo that is both phylogenetically and structurally coherent.
«Beyond the general requirement of monophyly there is no general criterion that dictates how inclusive a genus should be»
Still, by including this diverse array of fossils we are already beginning to strain at the limits of a morphologically coherent genus; and if we extend the erectus/ergaster group to embrace the amazing hominid finds from Dmanisi (Lordkipanidze et al., 2013) we find ourselves looking at a genus Homo that has been forced to accommodate within it an entirely implausible range of brain sizes and cranial morphologies. The same goes even more strongly for the <100,000 year old «Homo» floresiensis from Indonesia (Sutikna et al., 2016), and the South African Rising Star «Homo» naledi, now believed to date between about 335,000 and 236,000 years ago (Dirks et al., 2017). What is more, even having excluded the Dmanisi, Flores, and Rising Star hominids from Homo, we are still left with the array of African fossils in the range of about 1.6 million years or earlier that over the last four decades or so have been attributed to Homo habilis, H. rudolfensis, or simply to H. sapiens. Leakey et al. (2001) allocated rudolfensis to their new genus Kenyanthropus, while Collard and Wood (2015) sensibly excluded all these forms from Homo. Nevertheless, an overwhelming consensus still attributes all those ancient African fossils to our genus.
Figure 5. Clark Howell’s 1957 diagram of the temporal and spatial distributions of selected Middle and Late Pleistocene hominids. Note that Howell made no claim that this was a «tree». In blue, sites with human fossil sets. Redrawn by Kayla Younkin, adapted by Salvador Sastre. Figure 6. Highly tentative phylogeny of Hominidae as proposed by Ian Tattersall in 1993. / Drawn by Diana Salles, adapted by Salvador Sastre
When, deeply influenced by the notion of «man the toolmaker» (see Oakley, 1949), Louis Leakey and his colleagues gave the name Homo habilis to their 1.8 million years old new species from Tanzania’s Olduvai Gorge (Leakey, Tobias, & Napier, 1964), they did so within an established tradition of imputing fossil forms to our genus on the basis of inferred behaviors (meat-eating, language, toolmaking, art and symbolism, and so forth) rather than of observable morphology. But their action nonetheless unleashed a huge furor, in a science still in thrall to Mayr. Most observers were either unwilling to separate these fossils from an all-encompassing species Homo erectus, or refused to recognize any significant distinctions from Australopithecus specimens from South Africa. Proponents of the latter view had a particularly powerful point, for it seems clear that Leakey and colleagues’ principal reason for allocating the Olduvai fragments to a new, albeit more «primitive», species of Homo was that they were putatively associated with the crude stone tools also found in the lowest layer of the gorge.
Given the fact that any new behavior, even something as radical as the invention of stone tool making, must necessarily be invented by a member of an existing genus and species (who would hardly be expected to differ biologically in any significant way from his or her parents or offspring), Leakey and colleagues’ criterion for membership in the genus Homo was never very robust. Nonetheless, for a variety of (not very good) reasons, the notion of Homo habilis eventually caught on. In addition, a motley assortment of mostly very fragmentary fossils in the 2.0 to 2.5 million years ago range came to be attributed to «early Homo» – especially after the very earliest stone tools had been discovered at a site dated to more than 2.5 million years ago (Semaw et al., 1997).
«If future analyses and finds confirm that the hominids just mentioned are united by a common postcranial conformation, then we may find it best to recognize the genus ‘Homo’ not on cranial apomorphies, but on postcranial ones»
The dynamic initiated at Olduvai soon gave rise to a vigorous search for the «earliest Homo» that continues today, the latest and most ancient entrant being the 2.8 million years old Ledi-Geraru form from Ethiopia (Villmoare et al., 2015). Yet none of the «early Homo» pretenders has much morphological claim to belong to the genus that is defined by the highly derived Homo sapiens. Instead, what seems to be happening is that paleoanthropologists, while forced to recognize that there is diversity in the early hominid record, are nevertheless still trying to partition that diversity between the two established categories Australopithecus and Homo. And the desire to cram new morphologies into those two existing pigeonholes, at any cost, appears to be a lot stronger than any perception of the clear need to adjust the hominid systematic framework (at both the species and generic levels) to accommodate new morphologies.
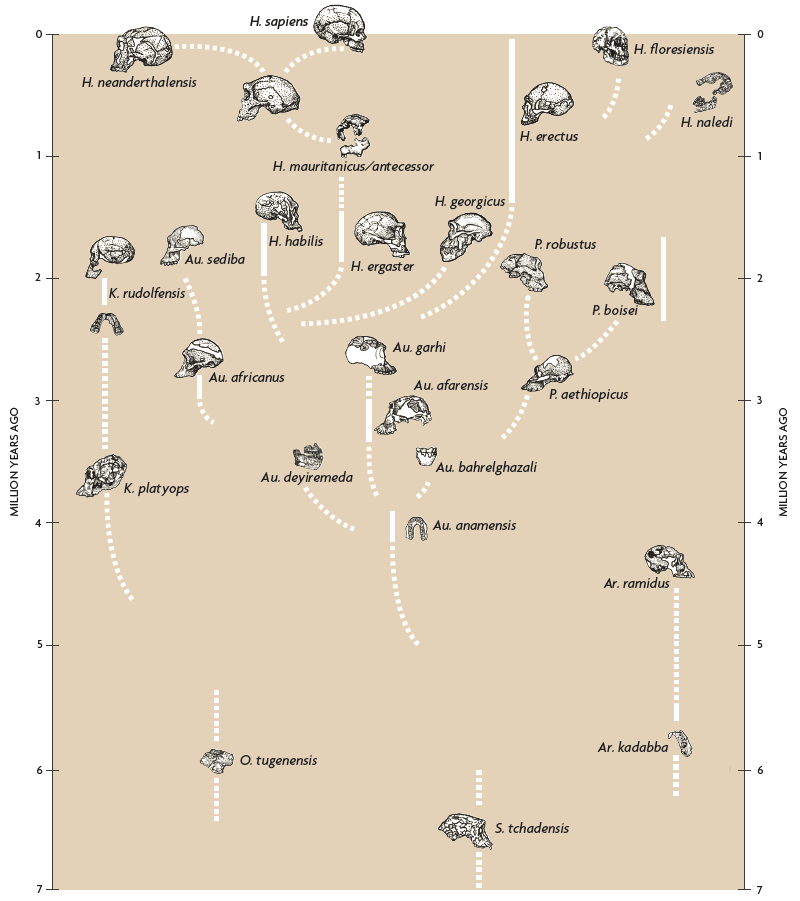
Figure 7. Highly tentative phylogeny of Hominidae as proposed by Ian Tattersall in 2017. / Drawn by Kayla Younkin, adapted by Salvador Sastre
Still, despite the heavy hand of tradition, it is plain by now that multiple lineages of hominids existed back into the distant past. And if we are to understand how this diversity was structured, it will surely be better to analyse it in terms of branching clades, rather than to cleave with increasing difficulty to the tradition of two dichotomised genera. Yes, indeed, genera are clades; but the morphological diversity that is emerging from the expanding hominid fossil record clearly needs to be reflected in a more expansive systematic context than tradition has permitted (Schwartz & Tattersall, 2015; Tattersall 2015b, 2016). Overstuffing a limited number of canonical genera will not ultimately be helpful in elucidating the evidently complex series of events that ultimately saw the emergence of Homo sapiens.
Conclusion
Throughout the history of palaeoanthropology the genus Homo has contracted and expanded, not in the service of phylogenetic and morphological coherence but according to philosophical fashion. Today finds us at an extreme of expansion, with a hugely ecumenical concept of a genus Homo that is both morphologically and phylogenetically untidy at best. At one rapidly-receding point there were signs that a very tentative reappraisal of the genus and its contents had begun, with the transfer of some iconic early Homo specimens to such genera as Australopithecus and Kenyanthropus (see Leakey et al., 2001; Collard & Wood, 2015). But the pendulum has since become stuck, with the result that we are still encumbered with a genus which is literally bursting at the seams, both at the morphological and systematic levels.
The lesson is surely that there is, or should be, nothing special about the genus Homo simply because Homo sapiens belongs to it. We should therefore abandon our hominid exceptionalism, and define our own genus exactly as other genera are defined. Achieving coherence in this way will involve a paring-down of the genus Homo, with an eye to recognizing a taxon that is firmly based on synapomorphy on the one hand, and is part of a more complex clade on the other that will require more genera to express its diversity. This will, of course, mean abandoning extravagant claims for the «earliest Homo» purely because the fossils concerned are not australopiths.
Acknowledgments
I thank Paquita Ciller and the late Jorge Wagensberg of CosmoCaixa (Barcelona, Spain) for their kind invitation to participate in the stimulating symposium on «The origin and evolution of the genus Homo» for which these remarks were prepared; the Fundación Caixa for making the symposium possible; my fellow participants for stimulating discussion; and Jordi Agustí for inviting me to publish this abbreviated version. Kayla Younkin kindly readied the illustrations for publication.
References
Arsuaga, J. L., Martínez, I., Arnold, L. J., Aranburu, A., Gracia-Téllez, A., Sharp, W. D., … Carbonell, E. (2014). Neandertal roots: Cranial and chronological evidence from Sima de los Huesos. Science, 344, 1358–1363. doi: 10.1126/science.1253958
Collard, M., & Wood, B. (2015). Defining the genus Homo. In W. Henke, & I. Tattersall (Eds.), Handbook of paleoanthropology (2n ed., vol. 3, pp. 2107–2144). Heidelberg: Springer.
Dirks, P. H. G. M., Roberts, E. M., Hilbert-Wolff, H., Kramers, J. D., Hawks, J., Dosseto, A., … Berger, L. R. (2017). The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. eLife, 6e24231. doi: 10.7554/eLife.24231
Dobzhansky, T. (1944). On species and races of living and fossil man. American Journal of Physical Anthropology, 2, 251–265. doi: 10.1002/ajpa.1330020303
Haeckel, E. (1868). Natürlische Schöpfungsgeschichte. Berlin: Reimer.
Howell, F. C. (1957). The evolutionary significance of variation and varieties of Neanderthal man. The Quarterly Review of Biology, 32, 330–347. doi: 10.1086/401978
Keith, A. (1915). The antiquity of man. London: Williams and Norgate.
King, W. (1864). The reputed fossil man of the Neanderthal. Quarterly Journal of Science, 1, 88–97.
Leakey, L. S. B., Tobias, P. V., & Napier, J. R. (1964). A new species of Homo from Olduvai Gorge. Nature, 202, 7–9. doi: 10.1038/202007a0
Leakey, M. G., Spoor, F., Brown, F. H., Gathogo, P. N., Leakey, L. N., & McDougall, I. (2001). New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature, 410, 433–440. doi: 10.1038/35068500
Linnaeus, C. (1758). Systema Naturae (10th ed.). Stockholm: Salvii.
Lordkipanidze, D., Ponce de León, M. S., Margvelashvili, A., Rak, Y., Rightmire, G. P., Vekua, A., & Zollikofer, C. P. E. (2013). A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science, 342, 326–331. doi: 10.1126/science.1238484
Mayr, E. (1950). Taxonomic categories in fossil hominids. Cold Spring Harbor Symposia on Quantitative Biology, 15, 109–118. doi: 10.1101/SQB.1950.015.01.013
Mayr, E. (1953). Comments on evolutionary literature. Evolution, 7(3), 273–281. doi: 10.2307/2405739
Oakley, K. P. (1949). Man the toolmaker. London: Trustees of the British Museum.
Robinson, J. T. (1951). Meganthropus, australopithecines and hominids. American Journal of Physical Anthropology, 11, 1–38. doi: 10.1002/ajpa.1330110112
Schwartz, J. H., & Tattersall, I. (2015). Defining the genus Homo. Science, 349, 931–932. doi: 10.1126/science.aac6182
Semaw, S., Renne, P., Harris, J. W. K., Feibel, C. S., Bernor, L., Fesseha, N., & Mowbray, K. (1997). 2.5 million-year-old stone tools from Gona, Ethiopia. Nature, 385, 333–336. doi: 10.1038/385333a0
Sutikna, T., Tocheri, M. W., Morwood, M. J., Saptomo, E., Jatmiko, Awe, R. D., … Roberts, R. G. (2016). Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia. Nature, 532, 366–369. doi: 10.1038/nature17179
Tattersall, I. (1993). The human odyssey. Englewood Cliffs, NJ: Prentice-Hall.
Tattersall, I. (2015a). Homo ergaster and its contemporaries. In W. Henke, & I. Tattersall (Eds.), Handbook of paleoanthropology (2nd ed., Vol. 3, pp. 2167–2188). Heidelberg: Springer.
Tattersall, I. (2015b). Defining and recognizing the genus Homo. Gortania, 36, 5–22.
Tattersall, I. (2016). The genus Homo. Inference: International Review of Science, 2(1). Retrieved from http://inference-review.com/article/the-genus-homo
Theunissen, B. (1988). Eugéne Dubois and the Ape-Man from Java: The history of the first “missing link” and its discoverer. Dordrecht/Boston: Kluwer Academic.
Villmoare, B., Kimbel, W. H., Seyoum, C., Campisano, E. N., DiMaggio, E. N., Rowan, J., … Reed, K. E. (2015). Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia. Science, 347, 1352–1355. doi: 10.1126/science.aaa1343
Weidenreich, F. (1939). Six lectures on Sinanthropus pekinensis and related problems. Journal of the Geological Society of China, 19, 1–110. doi: 10.1111/j.1755-6724.1939.mp19001008.x
Wood, B., & Collard, M. (1999). The human genus. Science, 284, 65–71. doi: 10.1111/j.1755-6724.1939.mp19001008.x