

Shedding New Light on How the Brain Works
Aportacions de les tècniques de neuroimatge a la neurociència cognitiva
Disseny experimental en neuroimatge: un exemple amb fMRI
| The growing development of functional neuroimaging techniques has involved a great advance in the study of the neural basis of the cognitive process. In this sense, these powerful techniques are helping to give an answer to questions such as where and when certain processes take place in the brain. However, it is a good experimental design that allows us to interpret the results given by these techniques according to contrasted theoretical models, which gives rise to answering the most important question of all: how is behaviour produced? | |||||||||||
|
The human being is curious by nature. He wants, even needs, to know how everything that surrounds him works. From childhood we are awed when we dismantle a device, we see what is inside and wonder what this or that component is for. And what is more astonishing: the more exceptional the function of a device is, the more we need to know about it. This simile can be applied to our brain for its exceptional role in our existence and adaptation to an increasingly complex world. Who has not sometimes wondered about the impressive brain functions? Why do we think, why do we have feelings as deep as love, compassion or rage, why are we creative, why…? But the most important thing is not so much knowing the why, but how the brain is capable of producing all these complex capabilities of the human being. Cognitive neuroscience is a relatively young discipline that since the nineties has been answering, at least partially, these questions, and formulating others; and for this aim it has made use of the potent techniques generically known as «functional neuroimaging techniques». The association between neuroimaging and cognitive neuroscience allows us to link brain activity and cognitive processes according to the following logic: the brain areas that show greater activation during the realization of a cognitive task are crucial for the cognitive function that requires said task. Thus, the combination of models that attempt to study the neural correlate of cognition and the data revealed by the neuroimaging techniques has exposed the existence of a large number of brain areas involved in any kind of processing. But the real challenge of cognitive neuroscience is to establish solid models of human cognition from which we can design experiments that give answer to how different interconnected brain areas work to produce behaviour. |
|||||||||||
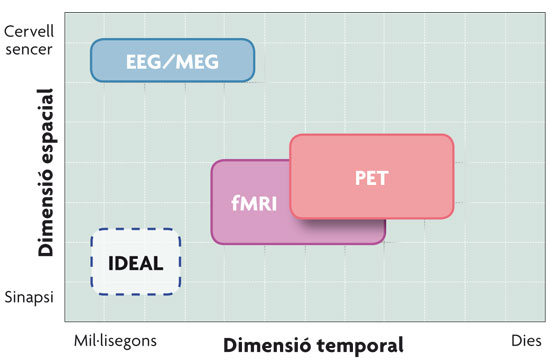
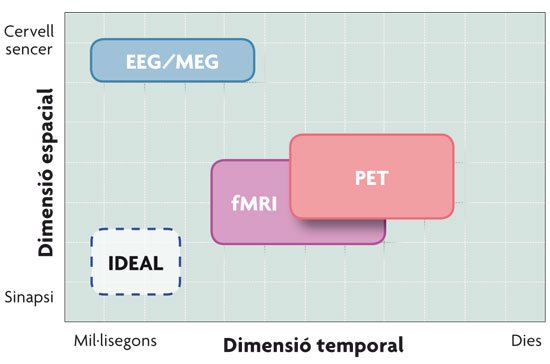 © Pilar Casado and Francisco Muñoz Comparison of the neuroimaging techniques described in the text in relation to their spatial and temporal resolution. As seen in the figure, the magnetoelectric techniques – EEG (electroencephalographic recording) and MEG (magnetoencephalography) – have better temporal resolution, while haemodynamic techniques – PET (Positron Emission Tomography) and fMRI (functional Magnetic Resonance Imaging) – have better spatial resolution. |
«Cognitive neuroscience is a relatively young discipline that studies how the brain is able to elaborate all the capabilities of the human being» |
||||||||||
|
A BRIEF OUTLINE OF NEUROIMAGING TECHNIQUES Functional neuroimaging techniques revolutionized cognitive neuroscience by allowing the study in real time of the physiological changes that occur in the brain of an individual while they perform a cognitive task (reading sentences, making decisions, watching images, etc.). In other words, cognitive neuroscience combines experimental strategies of cognitive psychology with various techniques dedicated to examine how brain function supports mental activity. The techniques that lead the current research about the working of the brain are: Event-Related Potentials (ERP), obtained from the electroencephalographic recording (EEG), magnetoencephalography (MEG), the Positron Emission Tomography (PET) and the functional Magnetic Resonance Imaging (fMRI). These four techniques enable us to record brain activity and are classified into two big groups, depending on the nature of the biological processes that said activity is associated with. Since the nerve impulse, the fundamental sign of neuronal communication, manifests at an electric, and therefore also magnetic, level, a first group of techniques of the cerebral activity record are the magnetoelectric techniques: EEG and MEG. These represent a direct measure of the neuronal activity, since they measure the electromagnetic fields generated by the flow of intracellular current produced in the cortical pyramidal neurons. Event-related potentials are variations of the basal magneto electroencephalographic register, related to the occurrence of an event and form a neurophysiological indicator of the processing underlying those stimuli. On the other hand, the neurons that are actively communicating need nutrients to function and, given their inability to store them, the brain depends on blood flow. Therefore, the regional increase in neural activity is associated with local increased metabolism and blood flow in the cerebral region. Haemodynamic techniques, mainly fMRI and PET, are responsible for measuring the activity of metabolic origin that occurs in certain parts of the brain as a result of neuronal activity; i.e., an area of the brain’s activity can be inferred from the amount of glucose and oxygen it consumes, substances that come through the bloodstream. The principle underlying both techniques is the same, that is, that the part (or parts) of the brain responsible for carrying out a certain cognitive task will increase their activity (and therefore the consumption of glucose and oxygen) in relation to the background activity. The fact that the processes that accompany these brain activations are different, as we have just seen, will also cause differences in the resolution of these techniques. The techniques that rely on cerebral haemodynamics have a very limited time resolution, since the increase in blood flow begins a few seconds after the onset of neuronal activity; but, on the other hand, the fact that there is not a single neuron in the brain not reached by the network of capillaries provides high spatial resolution to these techniques. Meanwhile, magnetoelectric techniques present a high temporal resolution, since they report on the brain changes millisecond to millisecond, but its spatial resolution is smaller, since they lack the capacity to report accurately on the activated brain areas. A solution to overcome this obstacle consists in combining techniques. For example, the combination of event-related potentials and fMRI permits to take advantage of the high temporal resolution of the first, and the high spatial resolution of the second. This advance has been possible thanks to the invention of non-ferromagnetic electrodes, which can be introduced in the strong magnetic field that is generated inside the fMRI scanner. |
|||||||||||
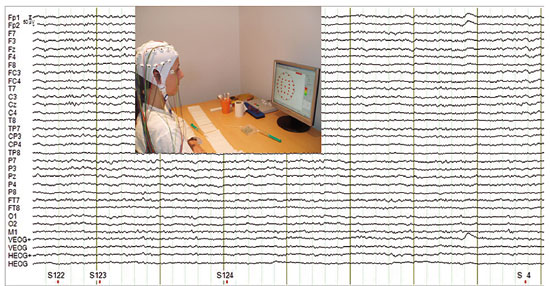 © Pilar Casado and Francisco Muñoz To perform an electroencephalographic recording it is necessary to place electrodes on the surface of the head, which will be in charge of detecting the underlying brain activity. In addition, an oculographic recording takes place, which allows to detect eye movements that may pollute the recording in order to remove them later. After getting the resistance provided by the different layers from the brain tissue (source of the activity that we want to record) to the surface (the place where we place the electrodes) down to acceptable levels (around 3KOhms), the recording of brain activity linked to the presented stimuli begins. |
«The real challenge of cognitive neuroscience is to establish solid models of human cognition from which we can design experiments that give answer to how different interconnected brain areas work to produce behaviour» |
||||||||||
|
THE IMPORTANCE OF DESIGN AND THE INTERPRETATION OF RESULTs As we have seen, brain-recording techniques provide us information about the areas of the brain that are activated during the realisation of various cognitive tasks, as well as of the temporality of the processes that are carried out. However, the mere knowledge of the location of the parts of an electronic device, for example, and the moment it starts up are useful to get an idea of how that appliance works, but do not provide a complete picture of its operation; it is necessary to know also the basic operating programme that manages the activity of the different components. This analogy applies to the study of brain function. It is clear that we have come a long way in the study of brain function thanks to the data that the development of advanced technologies has provided us with. But it is no less true that the interpretations and inferences made from said results, in order to get an idea about cognitive functioning, have not always been accurate. Well-known is the «study» by Craig Bennett, a neuroscientist specialized in the field of neuromarketing. He placed a dead salmon in a scanner and asked it to focus on the images that were on display and to try to identify the emotions on the faces of the people who appeared in them. The funny thing is that, by pure chance, there were signs of activity, specifically in parts of the salmon’s brain; obviously, there was no activity in the brain, but there were false positives, small measurement errors that sometimes affect some of the voxels, from which it could have been deduced that a dead salmon can identify human emotions. It is clear that we cannot draw conclusions from this study about the «cognitive functions» of the Atlantic salmon. Thus, it is essential to bear in mind that the data obtained with neuroimaging techniques has to be interpreted under the premises of at least two principles. Firstly, there must be a solid and verified theoretical model on which to base the cognitive function studies, which is to say that understanding the brain activity underlying a certain task has to have a theoretical framework as the starting point, which supports the results and allows to integrate them in a comprehensive manner. In addition, we must take into account that in neuroimaging experiments it is necessary to have a methodology and some very specific and limited experimental designs involving the isolation of different components of the cognitive processing, to best address the study. But this can lead to a misconception about the alleged «modular» and «encapsulated» character of cognition. There have been many advances in the study of the neural basis of cognitive functions made by following these guidelines. To name just one of them, which is particularly close to us authors, in the last years the neural basis of language models have been expanded. These were basically supported by studies of injuries that established that language is the product of the activity of a number of centres and connections between them, which essentially corresponded with Wernicke’s area, Broca’s area and their connection through the arcuate fasciculus. Thus, recent studies using fMRI on language processing establish that this is produced following a continuous flow of information, which is spread from the perisylvian region, primarily in the left hemisphere, to vast regions of the cerebral cortex (actually, nearly all cortical regions seem liable to participate). This includes for example the right hemisphere’s involvement in the processing of the prosodic aspects of language, or even the frontal motor areas in the semantic processing of words of action. On the other hand, the experimental paradigms that allow the use of the event-related potentials (ERP) technique for the study of linguistic anomalies, as is showed in the figure on this page, have permitted to identify a number of components of the ERP, indicators of the underlying processes, which are helping to clarify if the semantic and syntactic processing interact among themselves, or not, and if so, which one takes place first. |
|||||||||||
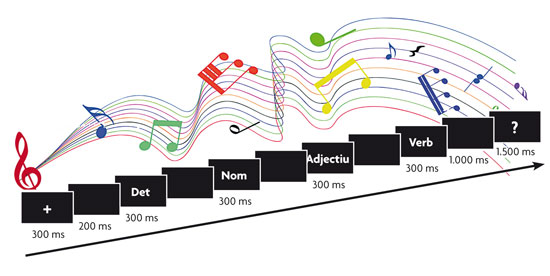 © Pilar Casado and Francisco Muñoz The experimental paradigm in the studies of sentence processing with potentials related to event-related potentials (ERP) usually involves the presentation of these sentences word-for-word, one of which has a syntactic or semantic anomaly and constitutes the stimulus target on which the event-related potential is analysed. In addition, we can include secondary tasks (listening to music in this example), to see how these affect the processing. |
«Brain-recording techniques provide us with information about the areas of the brain that are activated during the realisation of various cognitive tasks» |
||||||||||
|
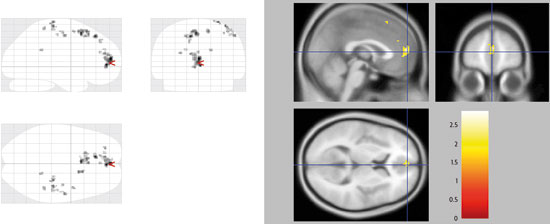
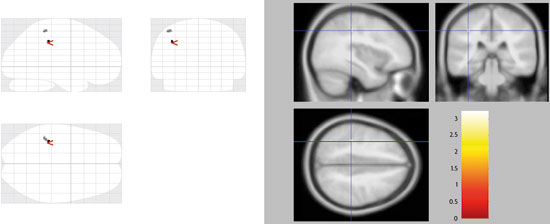
Besides, the combination of new brain imaging techniques and the most classic models allow us, without a doubt, to study language processing from a comparative perspective. The magnetic resonance imaging tractography is a novel method to study the orientation of the nerve tracts of white matter in vivo, i.e. it serves to explore the interconnections between different areas of brain function detected by fMRI. With this technique, it has been proven that the arcuate fasciculus seems to have evolved in the human brain from small tracts that connect the areas of Broca and Wernicke, already present in the macaque’s brain, more abundant in the chimpanzee’s, and with the maximum degree of development in humans. In this sense, one of the main differences between the brains of the human being and the chimpanzee is the remarkable expansion of the posterior ramifications of the arcuate fasciculus, particularly and notably within the left hemisphere, which has apparently played a key role in the evolution of human language. But this is just one example of how this new way to study how the brain works is being carried out; which has led to conducting experiments that allow us to study aspects unexplored until now, as are for example emotions and how they affect social relations. In recent studies by Professor Semir Zeki’s team it has been described that there is a single pattern of activation in the brain when we process the faces of people we hate, different from the pattern obtained when we process the faces of people we love, but there are common areas. This gives us an idea of why love and hate are so intertwined in our everyday life. In addition, when we feel love or hatred, certain cortical areas involved in reasoning and decision-making are disabled, and that is why we do not see the beloved or hated stimuli in a rational manner. Of all of the foregoing we can draw the conclusion that, in order to interpret the results correctly in the form of functional maps, we must know from the very beginning, as we are arguing, the theoretical model we use. By this, we mean that cognitive neuroscience should be aware of the limitations of the models and techniques it employs. In any case, we have to avoid falling into a new phrenology, trying to associate cognitive functions to a small set of specific brain areas, since there is increasingly more evidence in favour of the fact that these functions are possible because a complex network of brain areas is responsible for the neural processes that underlie said functions. In conclusion, the big questions of cognitive neuroscience regarding brain function have to be based on the integration of the theories about the processes that cognitive psychology provides and the correct application of the currently available and increasingly powerful techniques for the recording and analysis of brain activity, as well as those that could be developed in the future. REFERENCES Pilar Casado and Francisco Muñoz. Researchers of the Cognitive Neuroscience Section. Joint Centre for Research on Human Evolution and Behaviour. Professors at the Psychobiology Department. Complutense University of Madrid. |
|||||||||||
|
|||||||||||
BIBLIOGRAFIA
Bandettini, P. A., 2009. «What's New in Neuroimaging Methods?». Annals of the New York Academy of Sciences, 1.156: 260-293.
Bartels, A. i Zeki, S., 2000. «The Neural Basis of Romantic Love». Neuroreport, 11(17): 3.829-3.834.
Bennett, C. M. et. al., 2010. «Neural Correlates of Interspecies Perspective Taking in the Post-mortem Atlantic Salmon: an Argument for Multiple Comparisons Correction». Journal of Serendipitous and Unexpected Results, 1(1):1-5.
Buckner, R. L. i J. M. Logan, 2001. «Functional Neuroimaging Methods: PET and fMRI». In Cabeza, R. i A. Kingstone (eds.). Handbook of Functional Neuroimaging of Cognition. MIT Press. Cambridge.
Carretié, L., 2011. Anatomía de la mente. Emoción, cognición y cerebro. Pirámide. Madrid.
Kutas, M., et. al., 2006. «Psycholinguistics Electrified II: 1994-2005». In Traxler, M. i M. A. Gernsbacher (eds.). Handbook of Psycholinguistics. Elsevier. Nova York.
Martín-Loeches, M. P. i Casado, 2012. «Functional and Structural Magnetic Resonance Imaging of Human Language: A Review». In Bright, P. (ed.). Neuroimaging. Cognitive and Clinical Neuroscience. InTech. Rijeka.
Zeki, S. i J. P. Romaya, 2008. «Neural Correlates of Hate». PLoS One, 3(10): e3556.
Kawabata, H. i S. Zeki, 2004. «Neural Correlates of Beauty». Journal of Neurophysiology, 91: 1.699-1.705.




